







 и многоклеточных спорангиев органов бесполого размножения. Мужские половые органы называются антеридии, а женские архегонии. В антеридиях…")




 и получает от него питательные вещества. У зародыша папоротника, например, имеютс…")


. Отрасль систематики, изучающая мхи, называетс…")


 антеридии и архегонии. Обычно они располагаются группами, перемежаясь с многочисленными стерильными нитями парафизами, и окружены видоизмененными листовидными выростами.")







, представленного зеленым растением и бесполым (спорогоном) – развивающимся и живущим прямо на гаметофите.")
: 1. Антоцеротовые (Anthocerotopsida), 2. Печеночные (Marchantiopsida) и 3. Листостебельные (Bryopsida) мхи.")
 со стручковидной коробочкой.")

 поражают разнообразием форм гаметофитов, а спорофит имеет однотипное…")
















 видов и имеют первостепенное значение среди других мхов по участию в растительных сообществах различных зон.")








. Благодаря гиалиновым клеткам сфагновые мхи о…")


, в котором насчитывается свыше 300 видов. Во флоре России известно около 40 видов сфагновых мхов.")




 бриевые (Bryales) изобриевые (Isobryales) гипнобриевые (Hypnobryales) фунариевые (Funariales) гриммиевые (Grimmiales)")









")



Презентация на тему: Гаплоидная линия и моховидные

ВЫСШИЕ РАСТЕНИЯ Высшие растения подразделяются на две группы: высшие споровые и семенные растения. По сравнению с низшими растениями число видов высших очень велико и достигает по данным разных систематиков от 300 000 до 500 000.

Отделы высших растений К высшим споровым растениям относят отделы: моховидные, псилофитовидные, псилотовидные, плауновидные, хвощевидные и папоротниковидные. Семенные растения включают отделы: голосеменные и покрытосеменные.

Происхождение высших споровых Высшие растения произошли скорее всего от водорослей. Чередование поколений у высших споровых также аналогично чередованию поколений многих водорослей. Половые мужские клетки высших споровых имеют жгутики, так же как и гаметы или зооспоры водорослей. Имеется сходство в строении и функциях хлоропластов. Предками высших растений, вероятно, являлись зеленые пресноводные нитчатые или гетеротрихальные водоросли.

Эволюция высших растений

Эволюция жизненного цикла Эволюция жизненного цикла высших растений шла в двух противоположных направлениях. У моховидных она была направлена в сторону возрастания самостоятельности гаметофита и его постепенного морфологического расчленения, потере самостоятельности и редукции спорофита его морфологического упрощения. Остальные группы пошли по пути прогрессивного развития спорофита

Выход растений на сушу После выхода растений на сушу, который, вероятно, произошел в девоне, они попали в совершенно иную экологическую обстановку, отличную от условий жизни в воде и вынуждены были выработать специальные структуры, органы, ткани для сухопутной жизни.

Эпидерма и устьица Все органы спорофитов высших растений покрыты особым защитным слоем эпидермой, или кожицей. Для транспирации и газообмена с внешней средой в эпидерме в процессе эволюции возникли особые устройства –устьица, состоящие из 2-х замыкающих клеток

Особенности высших растений Для высших растений характерно наличие многоклеточных половых органов (гаметангиев) и многоклеточных спорангиев органов бесполого размножения. Мужские половые органы называются антеридии, а женские архегонии. В антеридиях развиваются мужские гаметы (сперматозоиды или спермии), а в архегониях женские гаметы (яйцеклетки).

Эволюция гаметангиев С усилением половой дифференциации количество мужских гамет в гаметангиях возрастало, а количество женских, наоборот, уменьшалось. Таким путем возник в конце концов архегоний с одной большой и неподвижной яйцеклеткой.

Вода в жизни споровых Чем ниже уровень эволюционного развития высших растений, тем в большей степени их половой процесс зависит от воды. У споровых растений оплодотворение яйцеклетки подвижными сперматозоидами происходит только при наличии воды, да и развитие самого гаметофита и даже спорофита нуждается в повышенной влажности.

Образование спор на спорофите На спорофите формируются органы бесполого размножения спорангии со спорами. Перед образованием спор в спорангиях у высших растений всегда происходит мейоз редукционное двухступенчатое деление, при котором из одной диплоидной материнской клетки образуются 4 гаплоидные споры. Споры имеют защитные оболочки – экзину и интину.

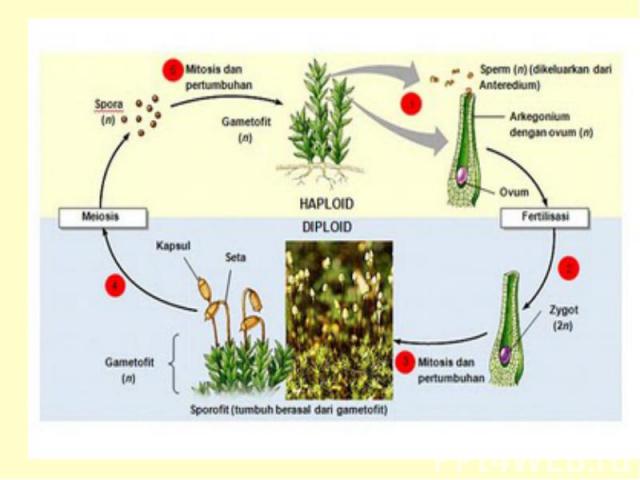
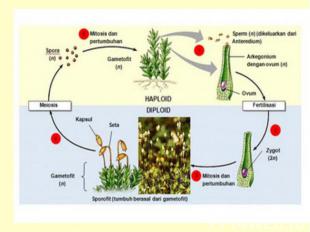
Жизненный цикл Гаплоидные споры прорастают в гаметофит – заросток На гаметофитах в гаметангиях формируются гаметы: сперматозоиды и яйцеклетки В результате оплодотворения, т.е. слияния мужской гаметы с женской, образуется зигота с двойным набором хромосом (отцовским и материнским). Зигота при прорастании дает начало многоклеточному зародышу, из которого развивается спорофит диплоидное поколение.

Зародыш споровых Из оплодотворенной яйцеклетки развивается зародыш. У современных споровых он имеет ножку, посредством которой внедряется в ткань заростка (гаметофита) и получает от него питательные вещества. У зародыша папоротника, например, имеются зачаточный корешок, стебелек и первый лист.

ПРЕДСТАВИТЕЛИ ГАПЛОИДНОЙ ЛИНИИ ЭВОЛЮЦИИ ВЫСШИХ СПОРОВЫХ РАСТЕНИЙ

Моховидные единственный отдел, представляющий гаплоидную линию эволюции высших растений. Основной жизненной формой у моховидных является гаплоидное поколение гаметофит, выполняющий кроме своей основной функции функцию фотосинтеза. Спорофит бесполое поколение сильно редуцирован и паразитирует на гаметофите, питаясь за его счет.

ОТДЕЛ BRYOPHYTA МОХОВИДНЫЕ Среди высших растений моховидные образуют обособленную группу. Собственно мхами систематики сейчас называют представителей листостебельных или настоящих мхов (класс Bryopsida). Отрасль систематики, изучающая мхи, называется бриологией. Древность моховидных подтверждается ископаемыми находками. В настоящее время известно примерно 23000 27000 видов моховидных.

Жизненный цикл мхов

гаметофит мхов помимо своей основной функции осуществлять половой процесс, выполняет и важные вегетативные функции фотосинтез, водное и минеральное питание.
 антеридии и архегонии. Обы")
На гаметофите формируются половые органы (гаметангии) антеридии и архегонии. Обычно они располагаются группами, перемежаясь с многочисленными стерильными нитями парафизами, и окружены видоизмененными листовидными выростами.

Архегонии представлены бутыльчатым многоклеточным образованием с узкой шейкой и брюшком, где помещается крупная яйцеклетка. При созревании архегония в шейке ослизняются особые канальцевые и шейковые клетки и по образующемуся каналу при наличии капельно-жидкой воды сперматозоиды могут проникнуть к яйцеклетке. Слияние гамет и дальнейшее развитие зиготы происходит внутри архегония.

Антеридии, архегонии и протонема

Жизненный цикл мхов Зигота прямо в архегонии прорастает в спорогон, на котором формируются органы бесполого размножения со спорами.

Протонема Спора прорастает в многоклеточное ветвистое или пластинчатое тело протонему, или предросток, похожее на водоросль. Протонема юношеская стадия развития гаметофита, характерная только для моховидных

Развитие протонемы На протонеме формируются новые многочисленные гаметофиты Протонема продолжает ветвиться, на ней закладываются все новые гаметофиты У некоторых мхов протонема является основной жизненной формой

Жизненный цикл мхов

Протонемы мхов

Чередование поколений Таким образом, у моховидных наблюдается чередование, или смена поколений: полового поколения (гаметофита), представленного зеленым растением и бесполым (спорогоном) – развивающимся и живущим прямо на гаметофите.


Разнообразие мхов На Земле произрастают около 25 000 видов моховидных. Этот отдел традиционно разделяют на три класса (отдела): 1. Антоцеротовые (Anthocerotopsida), 2. Печеночные (Marchantiopsida) и 3. Листостебельные (Bryopsida) мхи.

КЛАСС ANTHOCEROTOPSIDA АНТОЦЕРОТОВЫЕ В классе насчитывается около 300 видов. Центральное место принадлежит роду Anthoceros (около 200 видов) со стручковидной коробочкой.

Anthoceras

КЛАСС MARCHANTIOPSIDA , ИЛИ HEPATICOPSIDA ПЕЧЕНОЧНИКИ, ИЛИ ПЕЧЕНОЧНЫЕ МХИ КЛАСС MARCHANTIOPSIDA , ИЛИ HEPATICOPSIDA ПЕЧЕНОЧНИКИ, ИЛИ ПЕЧЕНОЧНЫЕ МХИ Печеночники (около 10 000 видов) поражают разнообразием форм гаметофитов, а спорофит имеет однотипное строение. У большинства протонема развита слабо и недолговечна. Среди печеночников есть талломные формы, но большинство имеет листостебельные побеги со спинно-брюшным расположением листьев.

Органы размножения печеночников
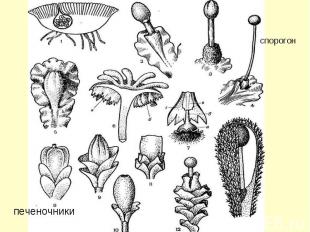

Вегетативное размножение

Подкласс маршанциевые Пор. Marchantiales Маршанциевые: Это талломные напочвенные мхи Marchantia polymorpha и Conocephalum Водный мох Riccia

Порядок Marchantiales Маршанциевые Маршанциевые обитатели влажных лесов или водные мхи. Наибольшего разнообразия они достигают в тропических и субтропических регионах, где среди них много эпифитов. В нашей флоре довольно часто можно встретить маршанцию многообразную (Marchantia polymorpha),

Подставки маршанции

Дихотомические талломы маршанций


Riccia

Conocephalum

ПОДКЛАСС JUNGERMANNIIDAE ЮНГЕРМАННИЕВЫЕ Порядок Metzgeriales Метцгериевые: Pellia, Blasia Порядок Jungermanniales Юнгерманние-вые: Plagiochila, Radula, Ptilidium

Pellia epiphylla

Pellia epiphylla - спорогон

Ptilidium

Radula

Plagiochila porelloides

КЛАСС BRYOPSIDA ЛИСТОСТЕБЕЛЬНЫЕ МХИ Листостебельные бриевые мхи включают наибольшее число (около 15000) видов и имеют первостепенное значение среди других мхов по участию в растительных сообществах различных зон.


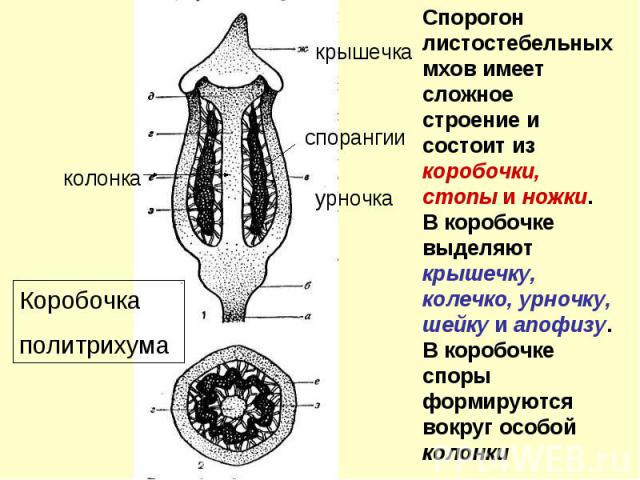
Положение гаметангиев и спорогонов У ортотропных мхов гаметангии и спорогоны развиваются на вершине стебля, таким образом мхи получаются верхоплодными. Плагиотропные мхи, наоборот, бокоплодны

Поперечный разрез стебля
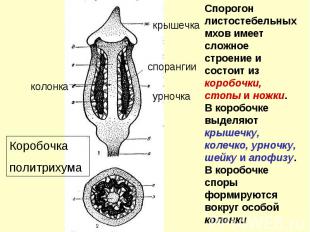

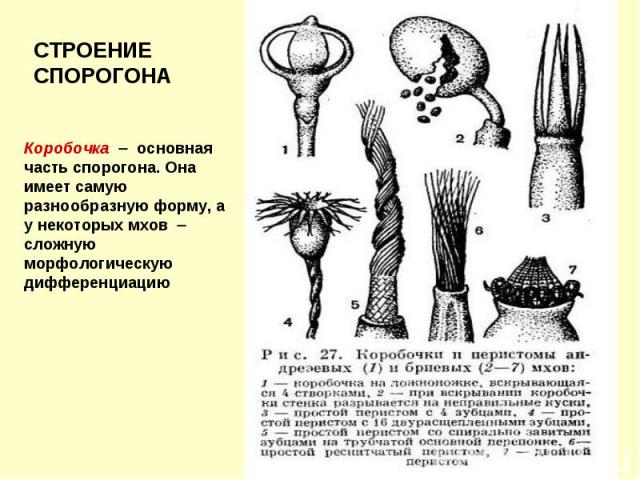
Коробочки бриевых мхов

Апофизы мхов Апофиза – расширенная часть коробочки Она часто окрашена

Перистом Для постепенного и своевременного рассеивания спор по краю урночки развит зубчатый гигроскопичный перистом

ПОДКЛАСС SPHAGNIDAE СФАГНОВЫЕ

Строение сфагновых мхов

Листья сфагновых мхов Все листья однослойные, без жилки и состоят из клеток двух типов узких хлорофиллоносных и широких бесцветных мертвых со спиральными утолщениями и порами (гиалиновых, или водоносных). Благодаря гиалиновым клеткам сфагновые мхи очень гигроскопичны и обладают огромной влагоемкостью


Строение стебля и листа сфагна

Разнообразие сфагновых В подклассе только один порядок Sphagnales с одним семейством сфагновых и одним родом сфагнум (Sphagnum), в котором насчитывается свыше 300 видов. Во флоре России известно около 40 видов сфагновых мхов.

ПОДКЛАСС ANDREAEIDAE АНДРЕЕВЫЕ Андреевые мхи являются мелкими, жесткими и хрупкими черно-бурыми мхами, растущими в виде подушечек на камнях и скалах.

Andreaea nivalis

Andreaea

Андреевые мхи Зрелая коробочка напоминает китайский фонарик и раскрывается 4 8 продольными щелями-створками,
 бриевые (Bryales)")
ПОДКЛАСС BRYIDAE БРИЕВЫЕ Порядки: политриховые (Polytrichales) бриевые (Bryales) изобриевые (Isobryales) гипнобриевые (Hypnobryales) фунариевые (Funariales) гриммиевые (Grimmiales)



Brachytecium






")
Наскальные мхи (Racomitrum )

Ptilium crista castrensis

Aulacomnium palustre

Происхождение мхов моховидные произошли непосредственно от водорослей (так называемая “водорослевая” теория происхождения











