


 ферменты, участвующие …")

 Гуанозинтрифосфат (ГТФ) Протеинкиназы АТФ.")
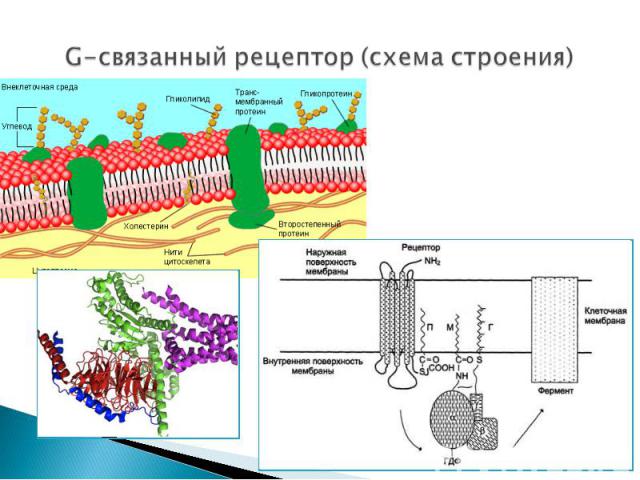
 с белком-рецептором происходят изменения конформации G-белка В результате одна из субъединиц G-белка (альфа) при…")
 с участием АТФ. В результате чего изменяется активность фосфорилированного белка - белки могут ста…")


, витамин D. Лиганды являются липоф…")

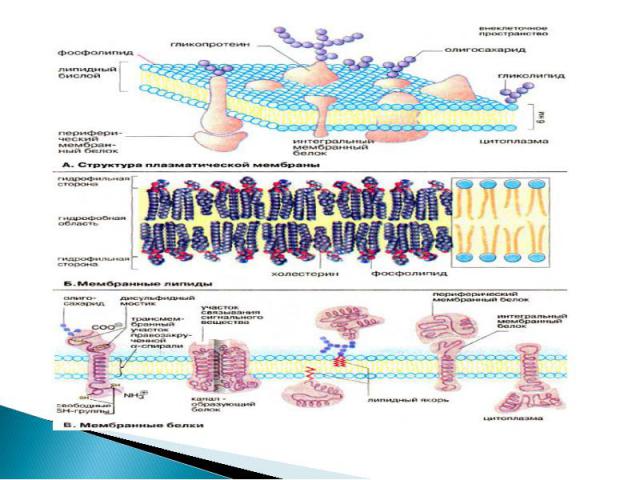
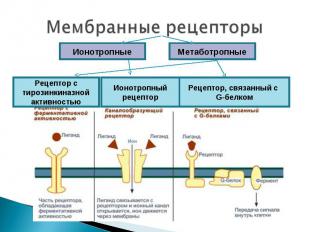
Презентация на тему: Рецепторы клетки



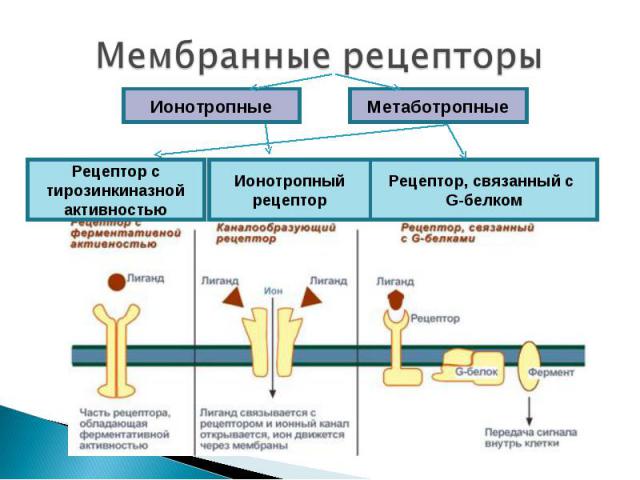
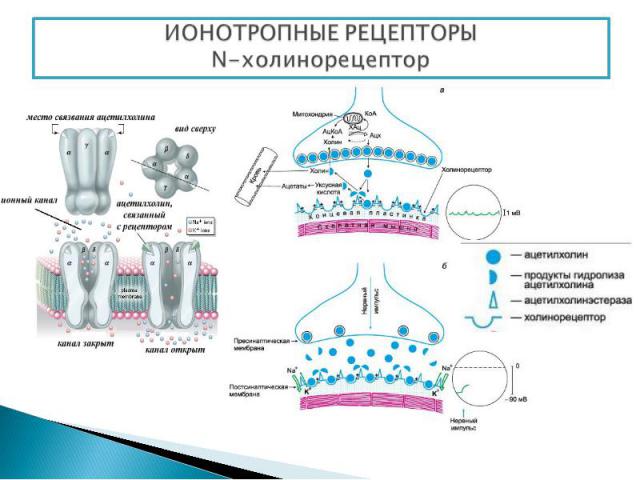
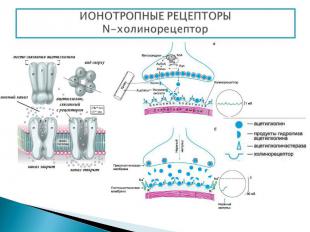
Представляют собой мембранные каналы, открываемые или закрываемые при связывании с лигандом, что приводит к ионному току через канал. N-холинорецептор - располагается в постсинаптической мембране нервно-мышечных синапсов Активация рецептора происходит при его взаимодействии с ацетилхолином. Ацетилхолин синтезируется в цитоплазме окончаний нейронов из холина и ацетилкоэнзима А (ацетил КоА) при участии цитоплазматического фермента холинацетилтрансферазы. Холин в свою очередь образуется в теле нейрона, ацетил Ко-А в митохондриях окончаний. Депонируется ацетилхолин в синаптических пузырьках (везикулах), в каждом из которых находится несколько тысяч молекул ацетилхолина. Структура: 5 интегральных белковых субъединиц (2α, β, γ, δ), которые окружают ионный натриевый канал. Взаимодействовать с ацетилхолином способны только α-субъединицы

Нервные импульсы вызывают высвобождение ацетилхолина в синаптическую щель, после чего он взаимодействует с холинорецепторами. Нервные импульсы вызывают высвобождение ацетилхолина в синаптическую щель, после чего он взаимодействует с холинорецепторами. Соединение ацетилхолина с N-холинорецептором приводит к открытию ионного канала. (Время пребывания канала в открытом состоянии составляет около 1 мс, и при этом через канал проходит примерно 50 000 ионов Na+). Резкое изменение концентрации Na+ приводит к развитию потенциала действия и мышца сокращается Ацетилхолин быстро гидролизуется ферментом ацетилхолинэстеразой. Образовавшийся при этом холин в значительном количестве (50%) захватывается пресинаптическими окончаниями, транспортируется в цитоплазму, где вновь используется для биосинтеза ацетилхолина. Остальная его часть поступает в кровь.


Участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях. Участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях. Виды «вторичных посредников»: циклические нуклеотиды (цАМФ и цГМФ) ферменты, участвующие в синтезе циклических нуклеотидов инозитолтрифосфат кальций-связывающий белок - кальмодулин, Ca 2+ протеинкиназы-ферменты фосфорилирования белков

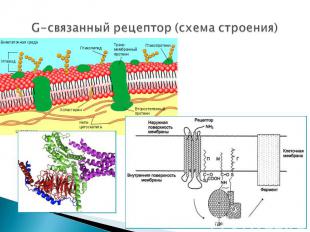
Представляют собой трансмембранные белки, имеющие 7 трансмембранных доменов, внеклеточный N-конец и внутриклеточный C-конец. Представляют собой трансмембранные белки, имеющие 7 трансмембранных доменов, внеклеточный N-конец и внутриклеточный C-конец. Сайт связывания с лигандом находится на внеклеточных петлях, домен связывания с G-белком — вблизи C-конца в цитоплазме. G-белок представляет собой гетеротример, состоящий из α , β, γ субъединиц. G-белок обладает ГТФ-азной активностью (способен гидролизовать ГТФ до ГДФ)

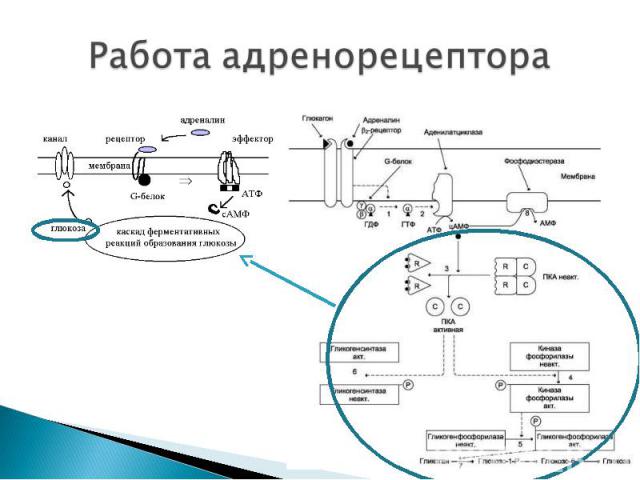
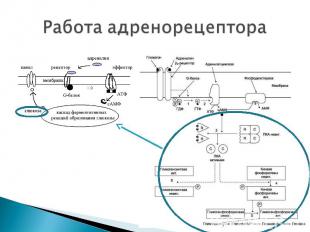
АДЕНИЛАТЦИКЛАЗНАЯ СИСТЕМА АДЕНИЛАТЦИКЛАЗНАЯ СИСТЕМА ФОСФОИНОЗИТИДНАЯ СИСТЕМА Компоненты аденилатциклазной системы: мембранный белок-рецептор G-белок Аденилатциклаза (фермент) Гуанозинтрифосфат (ГТФ) Протеинкиназы АТФ.

До момента действия гормона компоненты находятся в диссоциированнном состоянии. После образования комплекса сигнальной молекулы (гормон) с белком-рецептором происходят изменения конформации G-белка В результате одна из субъединиц G-белка (альфа) приобретает способность связываться с ГТФ ГДФ, связанная с белком обменивается на ГТФ G белок распадается на α- и β,γ-субъединицы. До момента действия гормона компоненты находятся в диссоциированнном состоянии. После образования комплекса сигнальной молекулы (гормон) с белком-рецептором происходят изменения конформации G-белка В результате одна из субъединиц G-белка (альфа) приобретает способность связываться с ГТФ ГДФ, связанная с белком обменивается на ГТФ G белок распадается на α- и β,γ-субъединицы. Комплекс “G-белок-ГТФ” активирует аденилатциклазу. Аденилатциклаза начинает активно превращать молекулы АТФ в ц-АМФ.

ц-АМФ активирует ферменты – протеинкиназы , которые катализируют реакции фосфорилирования различных белков (ферменты, ядерные белки, мембранные белки) с участием АТФ. В результате чего изменяется активность фосфорилированного белка - белки могут становиться функционально активными или неактивными. Такие процессы будут приводить к изменениям скорости биохимических процессов в клетке-мишени. ц-АМФ активирует ферменты – протеинкиназы , которые катализируют реакции фосфорилирования различных белков (ферменты, ядерные белки, мембранные белки) с участием АТФ. В результате чего изменяется активность фосфорилированного белка - белки могут становиться функционально активными или неактивными. Такие процессы будут приводить к изменениям скорости биохимических процессов в клетке-мишени. Активация аденилатциклазной системы длится очень короткое время. После гидролиза ГТФ до ГДФ альфа-субъединица отсоединяется от аденилатциклазы и G-белок восстанавливает свою конформацию (происходит реассоциация субъединиц) активация аденилатциклазы прекращается прекращается реакция образования цАМФ фермент – фосфодиэстераза катализирует реакцию гидролиза оставшихся ц-АМФ до АМФ. клеточный ответ прекращается

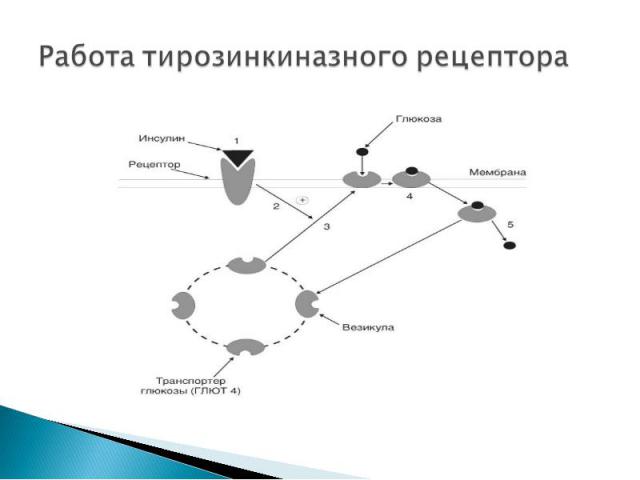
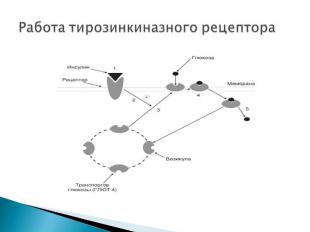
Структура: Структура: трансмембранные белки с одним мембранным доменом. Как правило, гомодимеры, альфа- и бета-субъединицы которых связаны дисульфидными мостиками. Лиганды: инсулин, факторы роста.

Инсулин с высокой специфичностью связывается и распознаётся α-субъединицей рецептора, которая при присоединении гормона изменяет свою конформацию. Инсулин с высокой специфичностью связывается и распознаётся α-субъединицей рецептора, которая при присоединении гормона изменяет свою конформацию. Это приводит к появлению тирозинкиназной активности у субъединицы β, что запускает разветвлённую цепь реакций по активации ферментов, которая начинается с аутофосфорилирования рецептора. Весь комплекс биохимических последствий взаимодействия инсулина и рецептора ещё до конца не вполне ясен, однако известно, что на промежуточном этапе происходит образование вторичных посредников (диацилглицеролов и инозитолтрифосфата) одним из эффектов которых является активация фермента — протеинкиназы С Протеинкиназа С обладает способностью фосфорилировать ферменты, что вызывает изменения во внутриклеточном обмене веществ. В результате происходит включение в клеточную мембрану цитоплазматических везикул, содержащих белок-переносчик глюкозы GLUT 4 GLUT 4 обеспечивает поступление глюкозы в клетку.

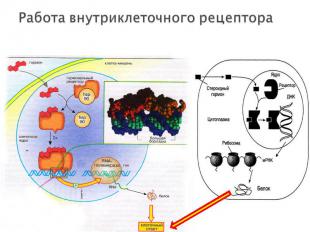
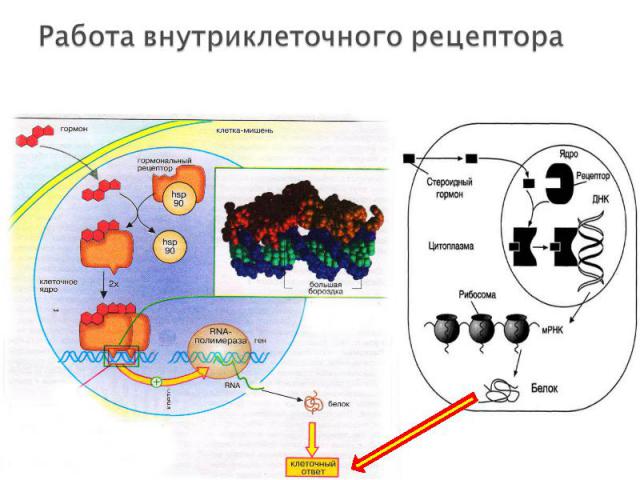
растворимые ДНК-связывающие белки растворимые ДНК-связывающие белки не связаны с плазматической мембраной, локализованы внутри ядра клетки!!! Лиганды: глюкокортикостероидные и тиреоидные гормоны (щитовидной железы), витамин D. Лиганды являются липофильными и способны проникать через мембрану внутрь клетки Структура рецептора: C-конец (гормон-связывающий домен) центральный домен (ДНК-связывающий) N-терминальный домен (активирующий транскрипцию гена-мишени)

проникновение гормона через билипидный слой мембраны в клетку проникновение гормона через билипидный слой мембраны в клетку образование комплекса гормон-рецептор, который перемещается в ядро клетки и взаимодействует с регуляторным участком ДНК. соответственно увеличивается (или уменьшается) скорость транскрипции структурных генов и скорость трансляции изменяется количество белков (в том числе ферментов), которые влияют на метаболизм и функциональное состояние клетки. Эффекты гормонов реализуются через определенный промежуток времени, так как на протекание матричных процессов (транскрипция и трансляция) требуется несколько часов. Возникший эффект продолжается еще некоторое время после удаления лиганда из комплекса.