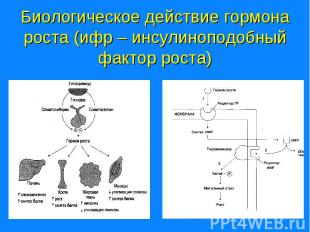
; Центральная нервная система (за счет передачи сигналов посредством нервных импульсов и нейромедиаторов); Эндокринная система (с помощью гормонов…")








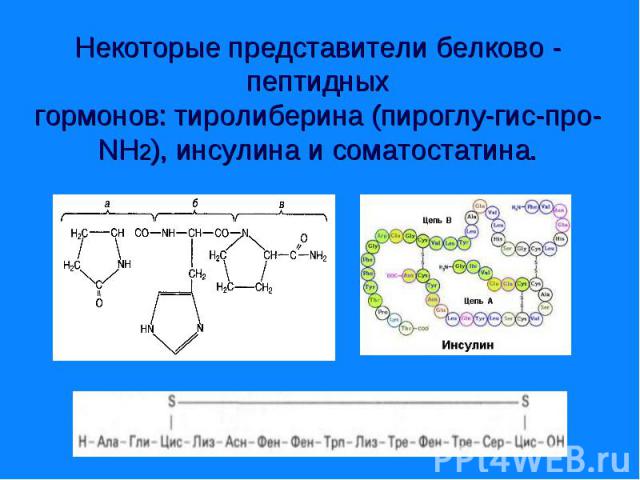
 и тироксин (I4), а также - адреналин и норадреналин – катехоламины.")
 Синтезируются из холестерина (на рис.) Гормоны коркового вещества надпочечников – кортикостероиды (кортизол, кортикостерон) Гормоны коркового вещества надпочечников – минералокортикоиды (андостерон) Половые гор…")
 — шир…")
 — синтезируются практически во всех клетках, кроме эритроцитов и лимфоцитов. Выделяют такие типы простагландинов A, B, C, D, E, F. Простагландины (Pg) — синтезируются практически во всех клетках, кроме эритроцитов и лим…")
 образуются в тромбоцитах, стимулируют их агрегацию и вызывают сужение мелких сосудов. Тромбоксаны (Tx) образуются в тромбоцитах, стимулируют их агрегацию и вызывают сужение мелких сосудов. Лейкотриены (Lt) активно синтезируются в ле…")



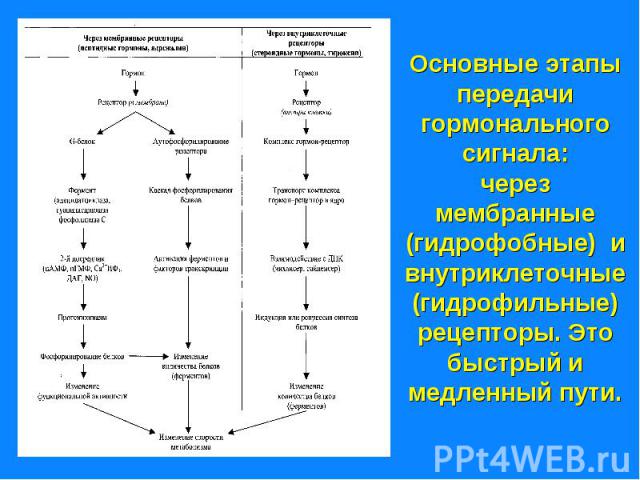



. Комплекс гормон-рецептор связан с G – белком, который имеет 3 субъединицы (α,β и γ). В отсутствии гормона α- субъединица связана с ГТФ и аденилатциклазой. Комплекс …")


 c G-белками (обозначены зеленым цветом). Рецепторы, сопряженные с G-белками, очень красивые, если рассматривать архитектурные молекулярные ансамбли клетки как шедевры природы. Взаимодействие б…")
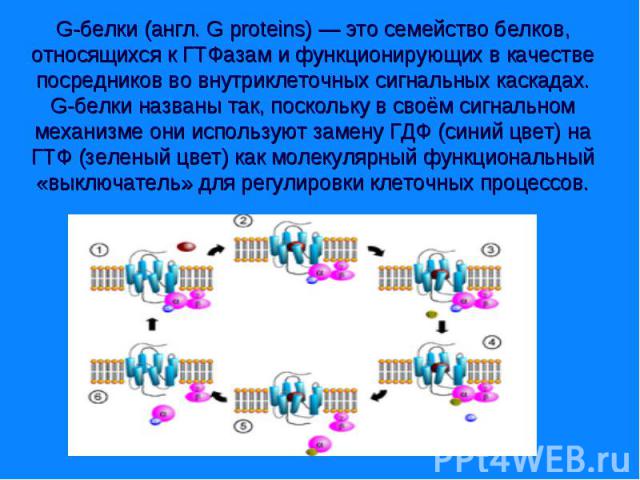
 и «малые». Гетеротримерные G-белки — это белки с четвертичной структурой, состоящие из трёх субъединиц: альфа(α), бета (β) и гамма (γ). Малые G-белки — это белки из одной полипепти…")


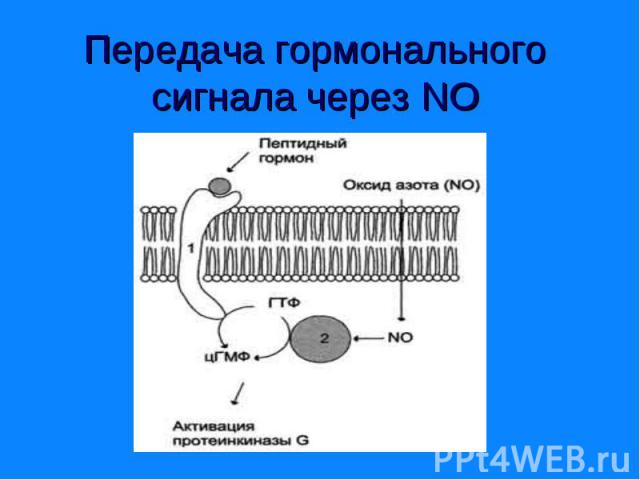
. Цитоплазматический домен рецептора обладает активностью гуанилатциклазы (гемсодержащий фермент). Молекулы цГТФ могут активировать ионные каналы или прот…")

Презентация на тему: ГОРМОНЫ

Автор – доцент Рыскина Е.А. Автор – доцент Рыскина Е.А.

Центральная нервная система (за счет передачи сигналов посредством нервных импульсов и нейромедиаторов); Центральная нервная система (за счет передачи сигналов посредством нервных импульсов и нейромедиаторов); Эндокринная система (с помощью гормонов, которые синтезируются в железах и транспортируются к клеткам-мишеням (на рис. А); Паракринная и аутокринная системы (при участии сигнальных молекул, секретируемых из клеток в межклеточное пространство - эйкозаноидов, гистаминов, гормонов ЖКТ, цитокинов) (на рис. Б и В); Иммунная система (посредством специфических белков – антител, Т-рецепторов, белков комплекса гистосовместимости.) Все уровни регуляции интегрированы и действуют как единое целое.


Дистантность действия – синтез в железах внутренней секреции, а регуляция отдаленных тканей Дистантность действия – синтез в железах внутренней секреции, а регуляция отдаленных тканей Избирательность действия Строгая специфичность действия Кратковременность действия Действуют в очень низких концентрациях, под контролем ЦНС и регуляция их действия осуществляется в большинстве случаев по типу обратной связи Действуют опосредованно через белковые рецепторы и ферментативные системы

Существует строгая иерархия или соподчиненность гормонов. Существует строгая иерархия или соподчиненность гормонов. Поддержание уровня гормонов в организме в большинстве случаев обеспечивает механизм отрицательной обратной связи.

Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет синтез гормонов, действуя либо на эндокринные железы, либо на гипоталамус. Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет синтез гормонов, действуя либо на эндокринные железы, либо на гипоталамус. Существуют эндокринные железы для которых отсутствует регуляция тропными гормонами – паращитовидная железа, мозговое вещество надпочечников, ренин-альдостероновая система и поджелудочная железа. Они контролируются нервными влияниями или концентрацией определенных веществ в крови.

по биологическим функциям; по биологическим функциям; по механизму действия; по химическому строению; различают 4 группы: 1. Белково-пептидные 2. Гормоны-производные аминокислот 3. Гормоны стероидной природы 4. Эйкозаноиды



Гормоны гипоталамуса; гормоны гипофиза; гормоны поджелудочной железы - инсулин, глюкагон; гормоны щитовидной и паращитовидной желез – соответственно кальцитонин и паратгормон. Гормоны гипоталамуса; гормоны гипофиза; гормоны поджелудочной железы - инсулин, глюкагон; гормоны щитовидной и паращитовидной желез – соответственно кальцитонин и паратгормон. Вырабатываются в основном путем прицельного протеолиза. У гормонов короткое время жизни, имеют от 3 до 250 АМК остатков.
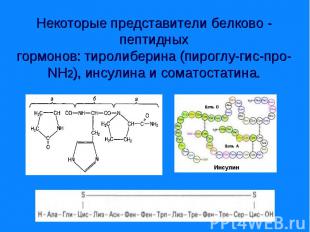

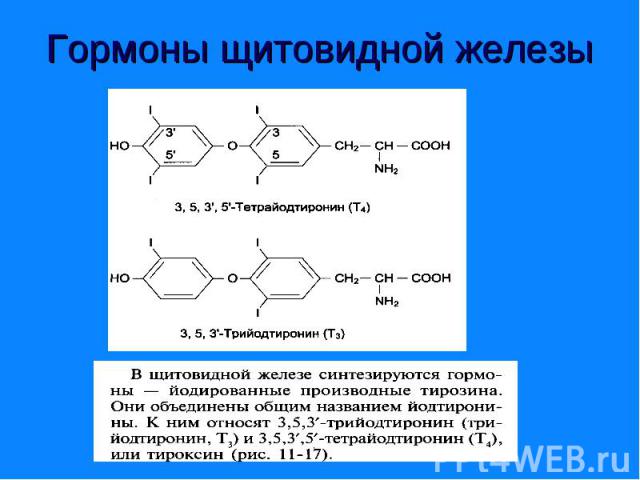
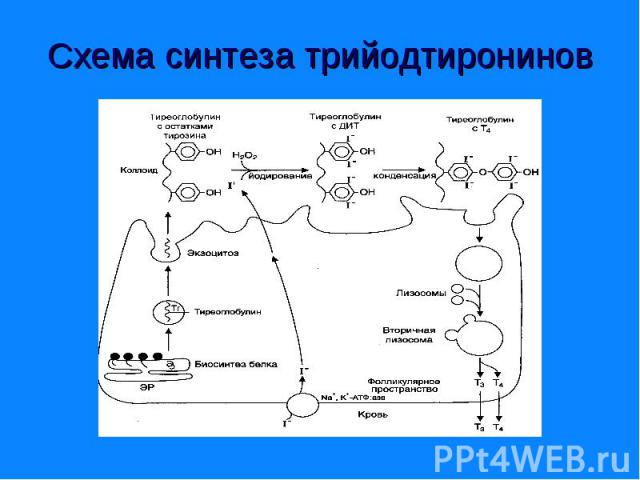
Являются производными аминокислоты - Являются производными аминокислоты - тирозина. К ним относятся гормоны щитовидной железы - трийодтиронин (I3) и тироксин (I4), а также - адреналин и норадреналин – катехоламины.

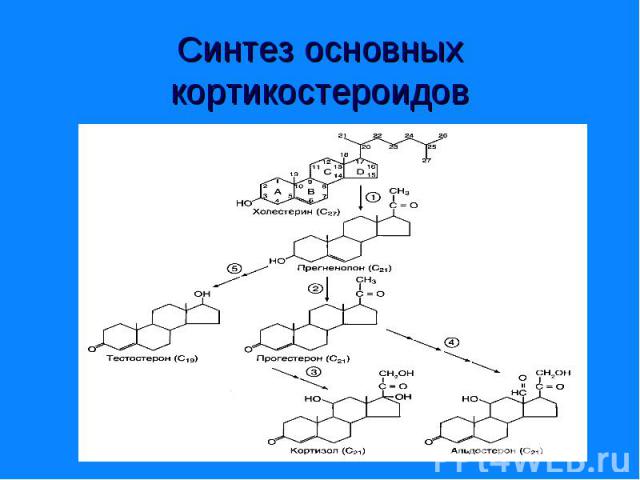
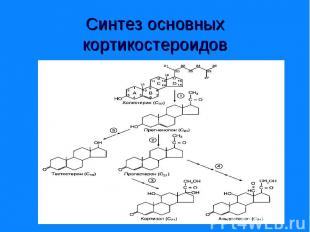
 Синтезируются из холестерина (на рис.) Го")
Синтезируются из холестерина (на рис.) Синтезируются из холестерина (на рис.) Гормоны коркового вещества надпочечников – кортикостероиды (кортизол, кортикостерон) Гормоны коркового вещества надпочечников – минералокортикоиды (андостерон) Половые гормоны: андрогены (19 «С») и эстрогены (18 «С»)

Предшественником всех эйкозаноидов является арахидоновая кислота. Предшественником всех эйкозаноидов является арахидоновая кислота. Они делятся на 3 группы – простагландины, лейкотриены, тромбоксаны. Эйказоноиды - медиаторы (локальные гормоны) — широко распространенная группа сигнальных веществ, которые образуются почти во всех клетках организма и имеют небольшую дальность действия. Этим они отличаются от классических гормонов, синтезирующихся в специальных клетках желез внутренней секреции.
 — синтезируются практически во всех клетках, кроме эрит")
Простагландины (Pg) — синтезируются практически во всех клетках, кроме эритроцитов и лимфоцитов. Выделяют такие типы простагландинов A, B, C, D, E, F. Простагландины (Pg) — синтезируются практически во всех клетках, кроме эритроцитов и лимфоцитов. Выделяют такие типы простагландинов A, B, C, D, E, F. Функции простагландинов сводятся к изменению тонуса гладких мышц бронхов, мочеполовой и сосудистой систем, желудочно-кишечного тракта, при этом направленность изменений различна в зависимости от типа простагландинов и условий. Они также влияют на температуру тела. Простациклины являются подвидом простагландинов (Pg I), но дополнительно обладают особой функцией — ингибируют агрегацию тромбоцитов и обусловливают вазодилатацию. Особенно активно синтезируются в эндотелии сосудов миокарда, матки, слизистой желудка.
 образуются в тромбоцитах, стимулируют их агрегацию и вызывают с")
Тромбоксаны (Tx) образуются в тромбоцитах, стимулируют их агрегацию и вызывают сужение мелких сосудов. Тромбоксаны (Tx) образуются в тромбоцитах, стимулируют их агрегацию и вызывают сужение мелких сосудов. Лейкотриены (Lt) активно синтезируются в лейкоцитах, в клетках лёгких, селезёнки, мозга, сердца. Выделяют 6 типов лейкотриенов: A, B, C, D, E, F. В лейкоцитах они стимулируют подвижность, хемотаксис и миграцию клеток в очаг воспаления. Также вызывают сокращение мускулатуры бронхов в дозах в 100—1000 раз меньших, чем гистамин.

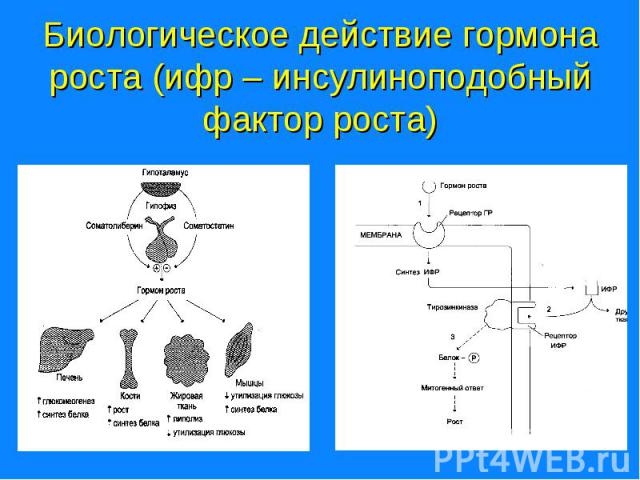
Для проявления биологической активности связывание гормонов с рецепторами должно приводить к образованию сигнала, который вызывает биологический ответ. Для проявления биологической активности связывание гормонов с рецепторами должно приводить к образованию сигнала, который вызывает биологический ответ. Например: щитовидная железа – мишень для тиротропина, под действием которого увеличивается количество ацинарных клеток, повышается скорость синтеза тиреоидных гормонов. Клетки-мишени отличают соответсвующий гормон, благодаря наличию соответствующего рецептора.


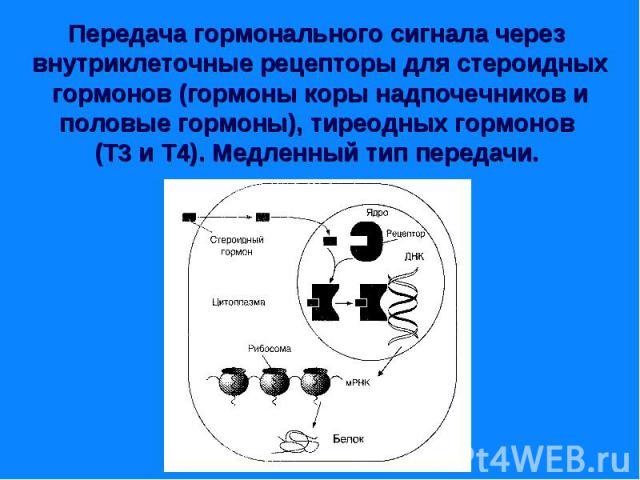
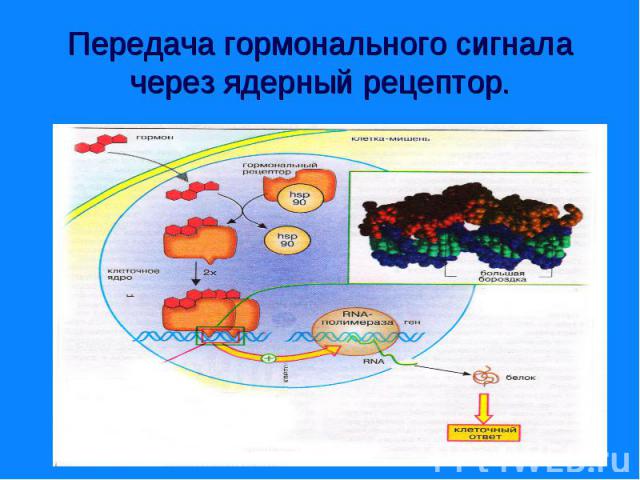
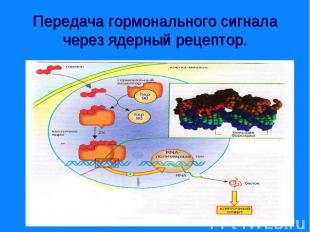
Рецепторы могут находится: Рецепторы могут находится: - на поверхности клеточной мембраны - внутри клетки – в цитозоле или в ядре. Рецепторы – это белки, могут состоять из нескольких доменов. Мембранные рецепторы имеют домен узнавания и связывания с гормоном, трансмембранный и цитоплазматический домены. Внутриклеточные (ядерные) – домены связывания с гормоном, с ДНК и с белками, регулирующие трансдукцию.
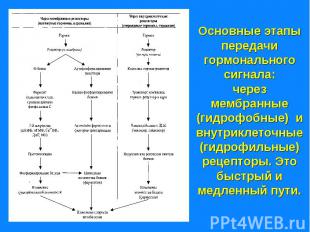

Гормональный сигнал меняет скорость метаболических процессов ответ путем: - изменение активности ферментов - изменение количества ферментов. Гормональный сигнал меняет скорость метаболических процессов ответ путем: - изменение активности ферментов - изменение количества ферментов. По механизму действия различают гормоны: - взаимодействующие с мембранными рецепторами (пептидные гормоны, адреналин, эйкозаноиды) и - взаимодействующие с внутриклеточными рецепторами (стероидные и тиреодные гормоны)
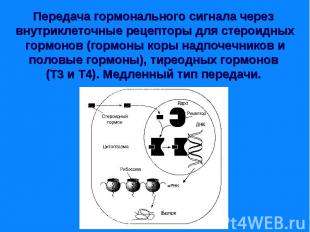

Передача информации от первичного посредника гормона осуществляется через рецептор. Передача информации от первичного посредника гормона осуществляется через рецептор. Этот сигнал рецепторы трансформируют в изменение концентрации вторичных посредников, получивших название вторичных мессенджеров. Сопряжение рецептора с эффекторной системой осуществляется через G –белок. Общим механизмом, посредством которого реализуются биологические эффекты является процесс «фосфорилирования – дефосфорилирования ферментов» Существуют разные механизмы передачи гормонального сигналы через мембранные рецепторы – аденилатциклазная, гуанилатциклазная, инозитолфосфатная системы и другие.

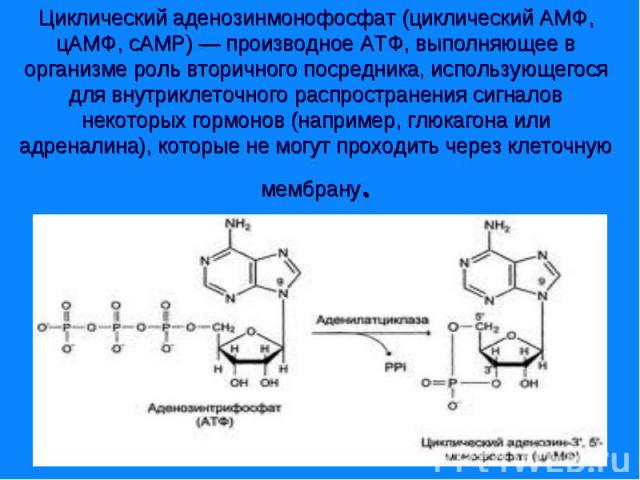
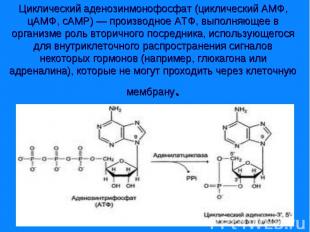
Сигнал от гормона трансформируется в изменении концентрации вторичных посредников – цАМФ, цГТФ, ИФ3, ДАГ, СА2+, NO. Сигнал от гормона трансформируется в изменении концентрации вторичных посредников – цАМФ, цГТФ, ИФ3, ДАГ, СА2+, NO.

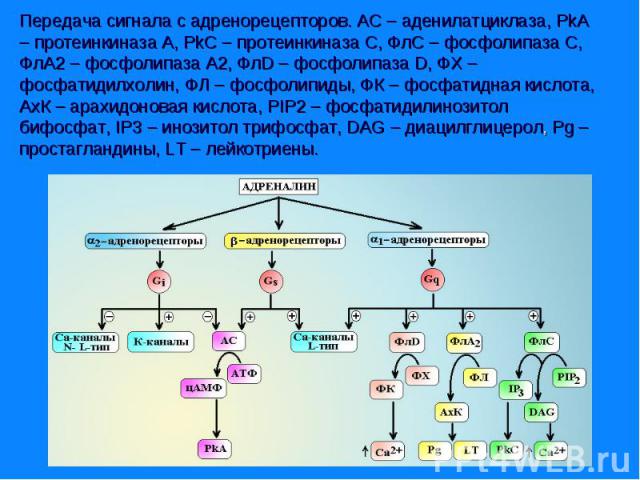
Комплекс гормон-рецептор связан с G – белком, который имеет 3 субъединицы (α,β и γ). Комплекс гормон-рецептор связан с G – белком, который имеет 3 субъединицы (α,β и γ). В отсутствии гормона α- субъединица связана с ГТФ и аденилатциклазой. Комплекс гормон-рецептор приводит к отщеплению димера βγ от αГТФ. Субъединица αГТФ активирует аденилатциклазу, катализирующую образование циклической АМФ (цАМФ). цАМФ активирует протеинкиназу А(ПКА), фосфорилируюшую ферменты, которые меняют скорость метаболических процессов. Протеинкиназы различают А,В,С и др.

Протеинкиназа А фосфорилирует ТАГ-липазу и активирует ее. Протеинкиназа А фосфорилирует ТАГ-липазу и активирует ее. ТАГ-липаза отщепляет от триацилглицеролов жирные кислоты с образованием глицерола. Жирные кислоты окисляются и обеспечивают организм энергией.
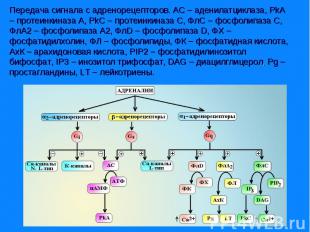

Адренорецепторы всех типов реализуют свое действие через Gs-белки. α- субъединицы этого белка активируют аденилатциклазу, которая обеспечивает синтез в клетке цАМФ из АТФ и активацию цАМФ зависимой протеинкиназы А. βγ-субъединицы Gs-белка активируют Са2+-каналы L-типа и макси-K+-каналы. Адренорецепторы всех типов реализуют свое действие через Gs-белки. α- субъединицы этого белка активируют аденилатциклазу, которая обеспечивает синтез в клетке цАМФ из АТФ и активацию цАМФ зависимой протеинкиназы А. βγ-субъединицы Gs-белка активируют Са2+-каналы L-типа и макси-K+-каналы. Под влиянием цАМФ-зависимой протеинкиназы А происходит фосфорилирование киназы легких цепей миозина и она переходит в неактивную форму, не способную фосфорилировать легкие цепи миозина. Процесс фосфорилирования легких цепей прекращается и гладкомышечная клетка расслабляется.
 c G-белками (обозначены")
Взаимодействие бета-2 рецептора (обозначен синим цветом) c G-белками (обозначены зеленым цветом). Рецепторы, сопряженные с G-белками, очень красивые, если рассматривать архитектурные молекулярные ансамбли клетки как шедевры природы. Взаимодействие бета-2 рецептора (обозначен синим цветом) c G-белками (обозначены зеленым цветом). Рецепторы, сопряженные с G-белками, очень красивые, если рассматривать архитектурные молекулярные ансамбли клетки как шедевры природы. Их называют «семиспиральными», поскольку они, спирально упакованы в клеточной мембране на манер елочного серпантина и «пронизывают» ее семь раз, выставляя на поверхность «хвостик», способный воспринять сигнал и передать конформационные изменения всей молекуле.
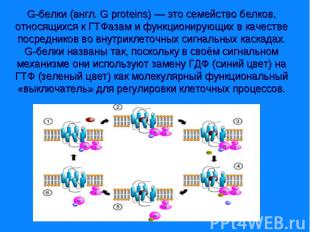
 и «малые».")
G-белки делятся на две основных группы — гетеротримерные («большие») и «малые». Гетеротримерные G-белки — это белки с четвертичной структурой, состоящие из трёх субъединиц: альфа(α), бета (β) и гамма (γ). Малые G-белки — это белки из одной полипептидной цепи, они имеют молекулярную массу 20—25 кДа и относятся к суперсемейству Ras малых ГТФаз. G-белки делятся на две основных группы — гетеротримерные («большие») и «малые». Гетеротримерные G-белки — это белки с четвертичной структурой, состоящие из трёх субъединиц: альфа(α), бета (β) и гамма (γ). Малые G-белки — это белки из одной полипептидной цепи, они имеют молекулярную массу 20—25 кДа и относятся к суперсемейству Ras малых ГТФаз. Их единственная полипептидная цепь гомологична α-субъединице гетеротримерных G-белков. Обе группы G-белков участвуют во внутриклеточной сигнализации.

Активность протеинкиназ типа А регулируется цАМФ, протеинкиназы G - цГМФ. Активность протеинкиназ типа А регулируется цАМФ, протеинкиназы G - цГМФ. Са2+ - кальмодулинзависимые протеинкиназы находятся под контролем концентрации СА2+. Протеинкиназы типа С регулируются ДАГ. Повышение уровня какого-либо вторичного посредника приводит к активации определенного класса протеинкиназ. Иногда субъединица мембранного рецептора может обладать активностью фермента. Например: тирозиновая протеинкиназа рецептора инсулина, активность которой регулируется гормоном.

Тирозинкиназа запускает процессы фосфорилирования внутриклеточных белков. Происходящее при этом аутофосфорилирование рецептора ведет к усилению первичного сигнала. Тирозинкиназа запускает процессы фосфорилирования внутриклеточных белков. Происходящее при этом аутофосфорилирование рецептора ведет к усилению первичного сигнала. Инсулин-рецепторный комплекс может вызывать активирование фосфолипазы С, образование вторичных посредников инозитолтрифосфата и диацилглицерола, активацию протеинкиназы С, ингибирование цАМФ. Участие нескольких систем вторичных посредников объясняет многообразие и различия эффектов инсулина в разных тканях.

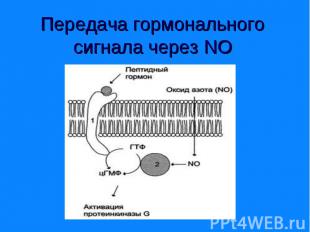
Цитоплазматический домен рецептора обладает активностью гуанилатциклазы (гемсодержащий фермент). Цитоплазматический домен рецептора обладает активностью гуанилатциклазы (гемсодержащий фермент). Молекулы цГТФ могут активировать ионные каналы или протеинкиназу G, фосфорилирующую ферменты. цГМФ контролирует обмен воды и ионный транспорт в почках и кишечнике, а в сердечной мышце служит сигналом релаксации.

Связывание гормона с рецептором, вызывает изменение конформациии рецептора. Связывание гормона с рецептором, вызывает изменение конформациии рецептора. Происходит диссоциация G-белка и ГДФ заменяется на ГТФ. Отделившаяся α-субъединица, связанная с молекулой ГТФ, приобретает сродство к фосфолипазе С. Под действием фосфолипазы-С происходит гидролиз липида мембраны фосфатидилинозитол-4,5-бисфосфата (ФИФ2) и образование инозитол-1,4,5-трифосфат (ИФ3) и диацилглицерол (ДАГ). ДАГ участвует в активации фермента протеинкиназы С (ПКС). Инозитол-1,4,5-трифосфат (ИФ3) связывается специфическими центрами Са2+-канала мембраны ЭР, это приводит к изменению конформации белка и открытию канала - Са2+ поступает в цитозоль. В отсутствие в цитозоле ИФ3 канал закрыт.