



 организована ретинотопически, причем максимальную поверхность занимает представительство центральной ямки (макулы), где плотность рецепторов максимальна. Площадь проекции …")



 зрение. Наиболее развито у приматов и человека в связи с фронтальным расположением глаз. Эта функция выполняется бинокулярными нейронами, активирую…")




 борозде и верхнем крае височной доли. Первичная слуховая кора (поле 41) организована тонотопически – нейроны, реагирующие на звуки разной частоты расположены …")

 и третичной (поле 22) имеют более…")

 и сенсорные (импрессивные) центры речи (рис.44). Повреждение моторных центров речи (зона Брока, 46 поле) вызывает моторную афазию Брока. При этом заболевании человека теряет способность говорит…")









 организованы соматотопически. Особенности нейронный …")






 функциям коры мозга человека относят: - сознание, - мышление, - речь, - научение и память, - внимание, - разработка стратегий адаптивного поведения.")
 получила дальнейшее развитие на современном этапе исследов…")
. Поле 6 – его повреждение вызывает аграфию (расстройство письма). Поле 44 – следствием его повреждения…")
, а поля 35 – …")


Презентация на тему: АНАЛИЗ ИНТЕГРАЛЬНОЙ ФУНКЦИИ

Анализаторные и интегративные функции новой коры

Анализаторные функции новой коры

Анализаторные функции новой коры. Зрение.
 ор")
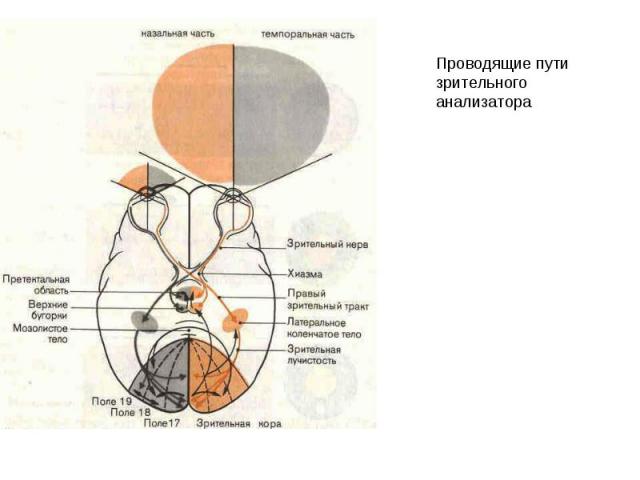
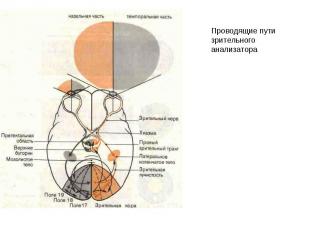
Анализаторные функции новой коры. Зрение. Первичная зрительная кора (поле 17) организована ретинотопически, причем максимальную поверхность занимает представительство центральной ямки (макулы), где плотность рецепторов максимальна. Площадь проекции периферического зрения зависит от плотности рецепторов на сетчатке. Нейроны первичной зрительной коры организованы в ориентационно-чувствительные и окулодоминантные колонки . Зрительная кора получает афферентную импульсацию по волокнам зрительной радиации от наружных коленчатых тел таламуса (НКТ) где заканчиваются волокна зрительного нерва, образованного аксонами ганглиозных клеток сетчатки. НКТ имеют 6-ти-слойное строение и проекции от правого и левого глаза поступают в соседние слои НКТ. Рецептивные поля нижних уровней зрительной системы (сетчатки, НКТ) круглые, в то время как рецептивные поля корковых нейронов имеют вытянутую форму и обладают ориентационной чувствительностью.

Ориентационные и окулодоминантные колонки зрительной коры

Простое рецептивное поле нейрона 17 поля зрительной коры

Нейроны со сложными и сверхсложными РП 18 и 19 полей зрительной коры

Бинокулярное зрение Одной из важных функций зрительной коры является бинокулярное (стереоскопическое) зрение. Наиболее развито у приматов и человека в связи с фронтальным расположением глаз. Эта функция выполняется бинокулярными нейронами, активирующимися при одновременной стимуляции идентичных (корреспондирующих) точек сетчатки правого и левого глаза. Бинокулярные нейроны обеспечивают объемное виденье предметов, за счет наличия клеток, способных измерять степень отклонения изображения предмета на сетчатке от корреспондирующих точек

Бинокулярное зрение

Цветовосприятие

Корковые механизмы зрительного опознания Конечным результатом зрительного восприятия является формирование зрительного образа объекта. Формирование зрительного образа и его опознание можно рассматривать как получение сжатого экономного описания объекта. У высших млекопитающих нейроны зрительной коры являются фильтрами пространственных частот, выделяющие такие признаки сигнала как форма, величина, цвет. У высших млекопитающих каналы, выделяющие определенные признаки объекта разделены функционально и морфологически. Так повреждение задневисочной коры (поле 37) вызывает у человека оптическую предметную агнозию – больной не может зрительно опознать объект, хотя узнает его на ощупь. При повреждении нижнетеменной области (поле 39) больной не может оценить пространственные отношения между предъявляемыми зрительными стимулами, определить их величину и направление движения.

Слуховая кора

Слуховая кора Первичная и вторичная слуховая кора располагается в латеральной (Сильвиевой) борозде и верхнем крае височной доли. Первичная слуховая кора (поле 41) организована тонотопически – нейроны, реагирующие на звуки разной частоты расположены в определенных локусах коры

Слуховая кора. Тонотопическая организация поля 41.

Слуховая кора Как и другие первичные проекционные зоны, слуховая кора имеет колончатую организацию. Нейроны одной колонки имеют одинаковые рецептивные поля и частотно-пороговую настройку. Нейроны вторичной (поле 42) и третичной (поле 22) имеют более обширные РП, по сравнению с нейронами первичной слуховой коры. У человека 22 поле – сенсорный центр речи Вернике, обеспечивающий понимание речи.

Центры речи
 и сенсорные (импрессивны")
Центры речи У человека выделяют моторные (экпрессивные) и сенсорные (импрессивные) центры речи (рис.44). Повреждение моторных центров речи (зона Брока, 46 поле) вызывает моторную афазию Брока. При этом заболевании человека теряет способность говорить, но продолжает понимать речь. Повреждение сенсорных центров речи (зона Вернике 22 поле) вызывает сенсорную афазию, при которой нарушается понимание речи. К речевым центрам относят также зоны, повреждение которых вызывает амнестическую афазию (поле 37) - забывание слов, алексию (поле 39) – расстройство чтения (речебразования), аграфию (поле 6) – расстройство письма, акалькулию – нарушение счета .

Центры речи

Слуховая кора Афферентные входы слуховая кора получает от медиальных коленчатых тел таламуса, в котором заканчиваются волокна латеральной петли, берущие начало от слуховых ядер продолговатого мозга и комплекса ядер верхних олив (латерального и медиального). Часть этих волокон заканчивается в задних буграх четверохолмия. Волокна слуховых путей многократно перекрещиваются, однако контралатеральные пути короче, чем ипсилатеральные. Медиальные коленчатые тела (МКТ) имеют ламинарную структуру, причем клетки каждого слоя иннервируются афферентными волокнами с двух сторон. Аксоны клеток МКТ образуют акустическую радиацию и оканчиваются на клетках слуховой коры. На всех уровнях слуховой системы имеются нисходящие пути, обеспечивающие обратные связи структур слуховой системы.

Проводящие пути слуховой коры

Слуховая кора На уровне коры анализ частоты звуков осуществляется по тонотопическому принципу. Интенсивность звука кодируется числом вовлеченных в реакцию нейронов и частотой их разрядов. Длительность звука кодируется медленноадаптирующимися тоническими нейронами слуховой коры. Для нейронов слуховой коры характерно привыкание (нейроны новизны) и усиление и стабилизация ответов на повторные раздражения (нейроны тождества) Важной характеристикой звукового сигнала является его частотная и амплитудная модуляции.

Соматосенсорная кора

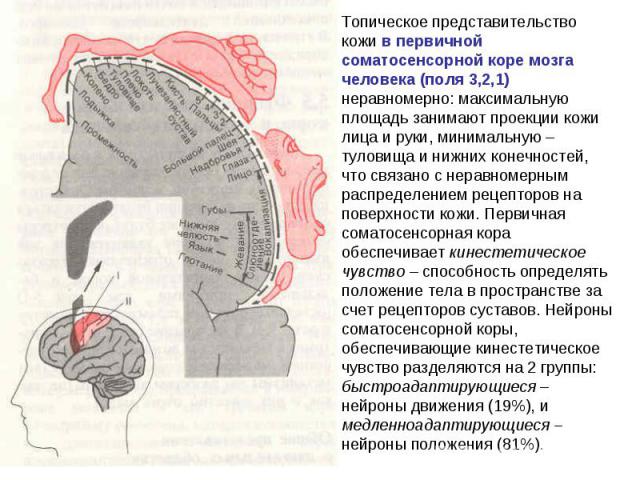
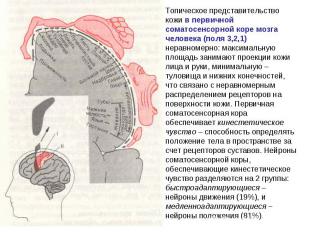
Соматосенсорная кора Соматосенсорная кора обеспечивает анализ информации, поступающей от рецепторов кожи, мышц и внутренних органов. Первичная соматосенсорная кора получает афферентную импульсацию от вентробазального комплекса ядер зрительного бугра, связанных с лемнисковыми путями - медиальным и тройничным лемнисками. Эта зона коры имеет типичную соматотопическую организацию.

Соматосенсорная кора Вторичная соматосенсорная зона коры получает информацию от задней группы ядер таламуса и не имеют топической организации. РП нейронов превышают по размеру поля первичной коры, а ответы нейронов имеют большую вариабельность по сравнению с ответами нейронов первичной зоны СМК. Во вторичной области соматосенсорной коры происходит анализ болевых раздражителей. В этой доне коры представлена в основном более древняя протопатическая болевая чувствительность, не позволяющая локализовать боль в отличие от эпикритеческой болевой чувствительности первичной соматосенсорной коры мозга.

Соматосенсорная кора Существует 2 пути проведения информации о температуре кожи в СМК от специфических температурных рецепторов: лемнисковый от холодовых рецепторов кожи по волокнам группы А, и спиноталамический путь проведения сигналов от тепловых и холодовых рецепторов кожи по волокнам группы С. Корковые нейроны реагирую в основном на перепады температур. В сенсомоторной коре представлен центральный отдел висцеральной системы.

Соматосенсорная кора Существует 2 пути проведения информации в структуры СМК лемнисковый от рецепторов кожи и мышц по волокнам группы А, и латеральный спиноталамический путь проведения сигналов от тепловых и холодовых рецепторов кожи В сенсомоторной коре представлен центральный отдел висцеральной системы.

Двигательная кора Функции двигательной коры впервые открыли Фритц и Гитциг в 1870 году, используя метод прямого электрического раздражения коры головного мозга. Двигательные зоны коры (поля 4 и 6) организованы соматотопически. Особенности нейронный организации двигательных корковых полей связаны с наличием гигантских пирамид Беца в пятом слое, а также наличие функциональных нейронных модулей, иннервирующих группу функционально однородных мотонейронов.

Двигательная кора

Двигательная кора Пирамидная система осуществляет контроль за проведением афферентных сигналов. При стимуляции пирамидного тракта наблюдается эффект подавления сенсорной импульсации. При этом возникают дорсальнокорешковые потенциалы, отражающие развитие пресинаптического торможения в задних корешках спинного мозга. Основная функция двигательных зон коры – управление тонкой моторикой и формирование программ двигательных актов. В двигательной коре хранится информация о двигательных паттернах, сформированных в течение всей жизни. В цепи выполнения движения двигательная коры выполняет функция первого звена, запускающего движение. Электрофизиологически это проявляется в формировании отрицательного потенциала, отражающего готовность к движению (Е-волна= волна ожидания).

Двигательная кора Повреждение двигательной коры вызывает паралич движений на стороне, противоположной повреждению, а также снижение мышечного тонуса. При повреждении пирамидных путей возникает спастическая гемиплегия – повышение тонуса разгибателей ноги и сгибателей руки на стороне, противоположной повреждению. Поражение премоторной и заднетеменной коры ограничивает возможности к подготовке двигательных актов и вызывает у человека апраксию – нарушение сложных двигательных актов.

Двигательная кора Латеральный кортико-спинальный путь Монакова оканчивается либо непосредственно на мотонейронах, либо на интернейронах спинного мозга. Этот путь регулирует тонкие движения дистальных отделов конечностей и позные рефлексы проксимальных мышц конечностей. Кортико-спинальные пути, идущие в передних канатиках спинного мозга заканчиваются на интернейронах медиальной группы и участвуют в поддержании равновесия и позы тела. Кортико-бульбарный тракты заканчиваются в продолговатом мозге в моторных ядрах лицевого и подъязычного нервов. Эти пути организованы также как и путь Монакова.

Интегративные функции новой коры.

Интегративные функции новой коры. Интегративные функции новой коры выполняются ассоциативными зонами коры, наиболее развитыми у приматов и человека – таламо-париетальной и эволюционно наиболее молодой таламо-фронтальной ассоциативной системой.
 функциям")
Интегративные функции новой коры. К высшим интегративным (психическим) функциям коры мозга человека относят: - сознание, - мышление, - речь, - научение и память, - внимание, - разработка стратегий адаптивного поведения.

Интегративные функции новой коры. Динамическая теория локализации функций И.П. Павлова, в основе которой лежит системный подход (1 и 2 сигнальные системы, ядро и периферия анализатора и т.п) получила дальнейшее развитие на современном этапе исследований и привела к формированию теории системной динамической локализации функций в коре, разработанной трудами академиков П.К.Анохина, А.Р. Лурия, Н.П. Бехтеревой, И.С. Беритова, О.С. Адрианова и их учениками.

Интегративные функции новой коры. Лобные доли коры Поле 8 – его повреждение вызывает нарушения статической координации, алексию (нарушение чтения). Поле 6 – его повреждение вызывает аграфию (расстройство письма). Поле 44 – следствием его повреждения является моторная афазия Брока, апраксия – нарушение произвольных движений Повреждение полей 9, 45,46,47 вызывает полифагию, снижение общего тонуса, нарушение внимания, памяти, мышления и эмоциональные расстройства (слева – депрессию, справа – эйфорию, благодушие). Нарушение функций полей 10 и 11 сопровождается двигательными расстройствами: атаксией, тремором, адиадохокинехом. При повреждении нижней поверхности лобных долей развивается расстройство обоняния – аносмия. Затылочные доли коры Поражение полей 19,39 вызывает оптическую агнозию – человек видит. но не узнает предметы.

Интегративные функции новой коры. Височные доли коры Диффузные поражения височных долей вызывают нарушения памяти. Повреждение поля 22 – сенсорную афазию Вернике. Нарушение функций поля 37 вызывает амнестическую афазию (забывание слов), а поля 35 – область крючка – обонятельную агнозию (нарушение распознавания запахов). Теменные доли коры Поражение полей 5 и 7 вызывает нарушения глубокой чувсвтельности, способности различать массу и форму предметов при их ощупывании). Поражение поля 40 левого полушария вызывает 2-х стороннюю моторную апраксию (теряется способность совершать привычные действия). Повреждение 39 поля вызывает алексию (нарушение чтения), аграфию и акалькулию (расстройства письма и счета). Поражение этих же полей правого полушария вызывает аутотопагнозию – нарушение схемы тела.

Интегративные функции новой коры. Ассоциативные таламокортикальные системы – это своеобразные структуры, не принадлежащие к какой-то определенной сенсорной системе, но получающие информацию от нескольких сенсорных систем. Эти системы получают сигналы от ассоциативных специфических ядер таламуса в свою очередь получающих сигналы от релейных (переключающих) ядер таламуса. Таким образом, ассоциативные зоны коры больших полушарий, не связанные непосредственно с сенсорными путями и интегрирующие разномодальные сигналы, обладают возможностью осуществлять оценку биологической значимости сигнала и выделять полезную информацию для формирования двигательных команд. Оценка биологической значимости сигнала сводится к установлению связи между сенсорными и двигательными системами мозга, которая у низших млекопитающих осуществляет напрямую, а у высших опосредована ассоциативными системами мозга.

Интегративные функции новой коры. В качестве основных механизмов работы ассоциативных систем определяют следующие: 1. Механизм мультисенсорной конвергенции, обеспечивающий выделение биологически значимых раздражителей для формирования целенаправленного поведенческого акта. 2. Механизм пластических перестроек при гетеромодальных сенсорных воздействиях, которые проявляютяс в привыкании, сенситизации, либо в формировании ответов экстаполяциооного типа. 3. Механизм краткосрочного хранения следов интеграции, заключающийся в длительной внутрикорковой и ли таламокорковой реверберации импульсных потоков.











