, отнесенную к расстоянию AB между этими точками:")

. . Различие становится особенно ощутимым, если учесть, что эта разница поддерживается на …")























 лежит прежде всего работа ионных каналов. Их В основе многих физиологических процессов (передача электрических…")
 или сакситоксин, продуцируемый некоторыми планктонами. Необратимо блокируют Na - ионные каналы такие паралитические …")

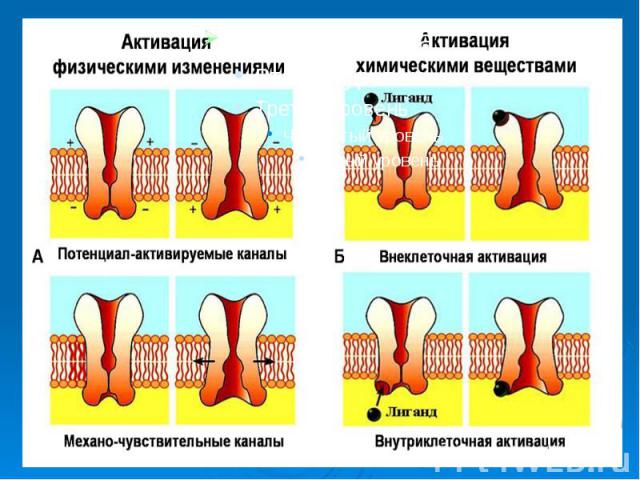
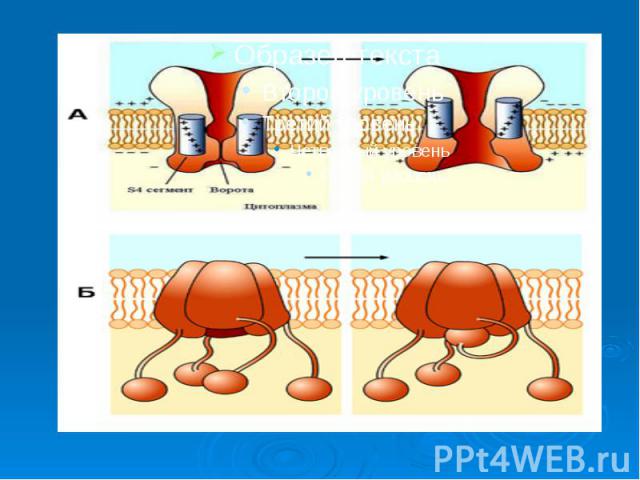
, а также способность открываться и закрываться при различных воздействиях на мембрану (воротная функция). Переходы каналов из открытого состояния в закрытое (воротн…")
. Эти каналы не имеют воротных механизмов, всегда открыты и почти не меняют свое состояние при электрических во…")




















. 3. Необходимость переносчика вещества (молекула - белковой природы). Присутствие фермента, который ак…")
. Для поддержания низко…")





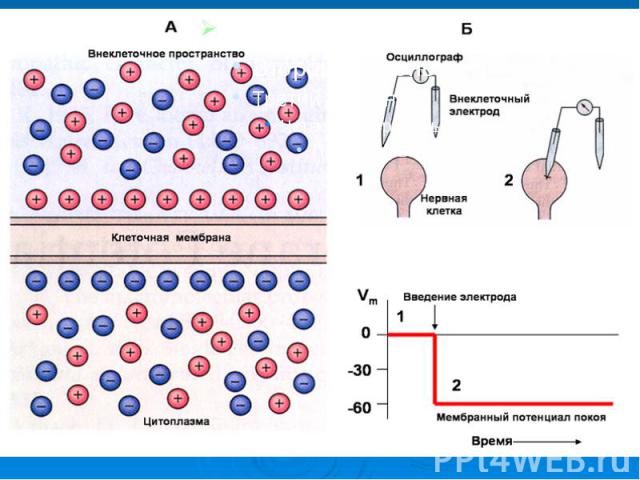
 и наружной поверхностями мембраны: Мембранным потенциалом называется разность потенциалов между внутренней (цитоплазматической) и наружной поверхностями мем…")
; выбором удачных…")
 представляет собой сте…")
, покрытая солью , …")
 - располагается в растворе (6) у наружной поверхности клетки (5) (рис. 11). Регистрирующее устройство осциллограф. Второй электрод сравнения (4) - располагается в растворе (6) у наружной поверхности клетки (5) (рис. 11)…")

 определяется трансмембранным градиентом к…")
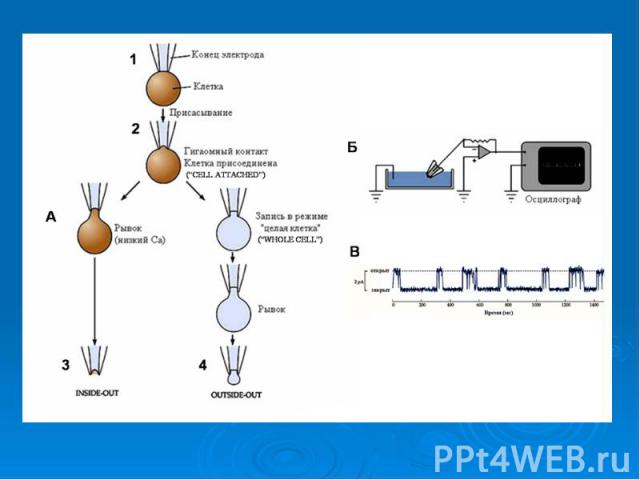
. Суть метода заключается в том, что микроэлектрод тонким концом, имеющим диаметр 0,5-1 мкм, присасывается к мембране таким об…")
. Сосуд, в котором находится растворы одной и той же соли, но разной концентрации, разделен пористой перегородкой (C1 > C2).")
. Вследствие разницы концентрации, ионы диффундируют, причем один из ионов, как правило, катион диффундирует быстрее и более разбавленный раствор приобретает знак катиона. По обе стороны перегородки образуется разность потенциалов, которую называют…")




 слушателям ВМА, в дальнейшем развил учение Ю.Бернштейн и лауреаты Нобелевской премии (Ходжкин, Кац, Хакси). Теория электролитиче…")
")















, или потенциал возбуждения нервных клеток (волокон), возникает в ответ на достаточное по силе раздражение. ПД - очень быстрый, кратковременный электрический процесс, поэтому для его регистрации необходим катодный осциллограф …")
 вводили на всю его длину два тончайших пр…")

, возникает потенциал действия . При усилении стимула и достижении порога раздражения, т.е. критического уровня деполяризации (КУД), возникает потенц…")

 и следовые потенциалы. В потенциале действия различают пик (спайк) и следовые потенциалы. Пик ПД представляет собой кратковременную инверсию (изменение знака на положительный) внутриклеточного потенциала. …")
 на 30 мВ. Эту разницу называют овершутом или амплитудой потенциала инверсии. Общая длительность пика в данном объекте составляет около 3м…")


, но сдвиг мембранного потенциала аксона ниже некоторого порогового значения ( критического уровня деполяризации –КУД )-возник…")
. Потенциал действия имеет стандартные амплитуду и временные параметры, не зависящие от силы…")


. Связь развития пика ПД с током доказывается прямой зависимостью амплитуды ПД от электрохимического градиента на мембране и достоверным переходом меченого изотопа из среды в клетку …")
 в нервной клетке. Этот ответ обладает в основном тем же механизмом, что и ПД. Его восходящая фаза определяется входящим током Na, а нисходящая - выходящим током . Рассмотрим локальный подпороговый ответ (…")











 и участками с меньшим потенциалом (отрицательно заряженными). Поэтому и в аксоплазме, и в…")





 от одного перехвата Ранвье (участка, свободного от миелиновой оболочки) до другого. Возбуждение по миелинизированному волокну распространяется сальтаторно (скачко…")

Презентация на тему: БИОФИЗИКА КЛЕТКИ

БИОФИЗИКА КЛЕТКИ Лекция 1. Транспорт веществ через биологические мембраны

План лекции 1. Введение 2. Градиенты биологических систем 3. Ультраструктура и функции биологических мембран 4. Пассивный транспорт 4.1. Диффузия 4.1.1. Транспорт ионов с помощью подвижного переносчика 4.1.2. Эстафетный перенос 4.1.3. Перенос ионов через каналы биомембран 4.2. Осмос 4.3. Фильтрация 4.4. Водный обмен между кровь и лимфой 5. Активный транспорт

1. Введение Структурной единицей живого организма является клетка, которой присущи все основные жизненные функции. Клетка - открытая термодинамическая система, которая обменивается с окружающей средой веществом, энергией и информацией.

Биофизика клетки является разделом науки, который связывает воедино биофизические исследования, проводимые на атомном и молекулярном уровне с исследованиями, проводимыми на уровне органов и биологических систем в целом. Биофизика клетки является разделом науки, который связывает воедино биофизические исследования, проводимые на атомном и молекулярном уровне с исследованиями, проводимыми на уровне органов и биологических систем в целом.

В середине 19 века великий французский физиолог Клод Бернар высказал гипотезу существования плазматической мембраны, до него биологи-морфологи представляли клетку в виде комочков живого вещества, не имеющих оболочки. В середине 19 века великий французский физиолог Клод Бернар высказал гипотезу существования плазматической мембраны, до него биологи-морфологи представляли клетку в виде комочков живого вещества, не имеющих оболочки.

. Выживание организма в условиях непрерывно меняющейся среды обусловлено способностью живых систем сохранять свое стационарное состояние. Для обеспечения такого состояния у всех организмов - от простых до самых сложных - существуют различные анатомические, физиологические и поведенческие приспособления. Все они направлены на поддержание постоянства внутренней среды. . Выживание организма в условиях непрерывно меняющейся среды обусловлено способностью живых систем сохранять свое стационарное состояние. Для обеспечения такого состояния у всех организмов - от простых до самых сложных - существуют различные анатомические, физиологические и поведенческие приспособления. Все они направлены на поддержание постоянства внутренней среды.

Положение о том, что именно постоянство внутренней среды определяет оптимальное условие для жизни и размножения организма было высказано К. Бернаром в 1857 году. Действительно, внутренняя среда высших животных обладает относительным постоянством физико-химических параметров. Для неё характерно постоянство температуры, рH, артериального давления, содержание кислорода, углекислого газа, различных ионов, сахаров и других физиологических констант, величины которых находятся в постоянной зависимости от структур биологической мембраны .

В 1929 году крупнейший американский физиолог Уолтер Б. Кэннон, расширив концепцию К. Бернара, создал учение о гомеостазе В 1929 году крупнейший американский физиолог Уолтер Б. Кэннон, расширив концепцию К. Бернара, создал учение о гомеостазе

. Гомеостаз - это универсальное свойство живых организмов активно сохранять стабильность работы всех систем организма в ответ на воздействия, нарушающие эту стабильность, и поддерживать динамическое постоянство внутренней среды, а также устойчивость основных физиологических функций организма. . Гомеостаз - это универсальное свойство живых организмов активно сохранять стабильность работы всех систем организма в ответ на воздействия, нарушающие эту стабильность, и поддерживать динамическое постоянство внутренней среды, а также устойчивость основных физиологических функций организма.

Это динамическое постоянство возможно только за счет непрерывного обмена веществом и энергией между внеклеточной и внутриклеточной средой - с одной стороны, Это динамическое постоянство возможно только за счет непрерывного обмена веществом и энергией между внеклеточной и внутриклеточной средой - с одной стороны, и между внеклеточной средой и внешней средой, окружающей органы и организм в целом - с другой стороны.

Такой обмен возможен лишь при наличии градиентов на границах перечисленных сред. . Такой обмен возможен лишь при наличии градиентов на границах перечисленных сред. .

2.Градиенты биологических систем

Под градиентом того или иного параметра системы для линейных функций подразумевают разность величин этого параметра X в двух точках пространства (A, B), отнесенную к расстоянию AB между этими точками:

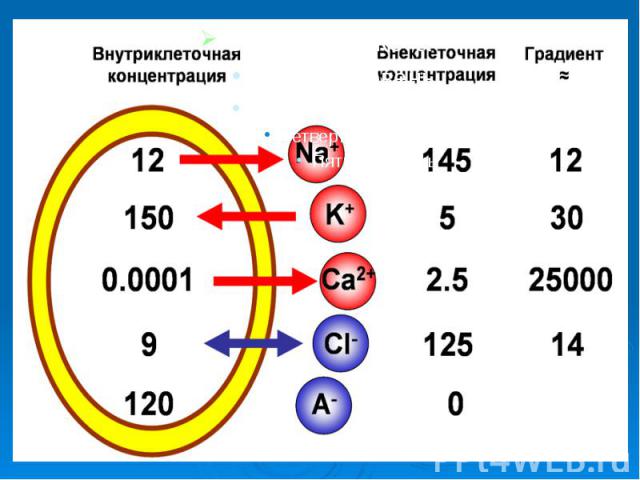
Заметим, что градиент - это вектор. Обилие градиентов является одним из отличий организма от неживых систем. Наиболее выражены градиенты на границе между цитоплазмой и внеклеточной средой. На первый взгляд это не столь уж большая разница в содержании тех или иных веществ в клетке и вне её. Например, в цитоплазме нервного волокна млекопитающих содержится 150 мМ/л, а в межклеточной жидкости - 5,0 мМ/л ионов калия. Разница - 145 мМ/л. Заметим, что градиент - это вектор. Обилие градиентов является одним из отличий организма от неживых систем. Наиболее выражены градиенты на границе между цитоплазмой и внеклеточной средой. На первый взгляд это не столь уж большая разница в содержании тех или иных веществ в клетке и вне её. Например, в цитоплазме нервного волокна млекопитающих содержится 150 мМ/л, а в межклеточной жидкости - 5,0 мМ/л ионов калия. Разница - 145 мМ/л.

. Различие становится особенно ощутимым, если учесть, что эта разница поддерживается на ничтожно малом расстоянии - порядка 10-8 м (толщина клеточной мембраны). . Различие становится особенно ощутимым, если учесть, что эта разница поддерживается на ничтожно малом расстоянии - порядка 10-8 м (толщина клеточной мембраны). Рассчитаем концентрационный градиент между клеточной мембраной по калию: пятнадцатимиллиардный градиент.


Столь же велики концентрационные градиенты для других ионов, белков, углеводов и т.д. Не менее выражены для клетки такие градиенты, как осмотический и электрический. Важно понять, что именно градиент, а не просто разность величин, является движущей силой многих жизненных процессов, в частности, переноса веществ между клетками и внеклеточной средой, а также между внутренней и внешними средами организма. Столь же велики концентрационные градиенты для других ионов, белков, углеводов и т.д. Не менее выражены для клетки такие градиенты, как осмотический и электрический. Важно понять, что именно градиент, а не просто разность величин, является движущей силой многих жизненных процессов, в частности, переноса веществ между клетками и внеклеточной средой, а также между внутренней и внешними средами организма.

Многие патологические процессы, связанные с нарушением транспорта веществ в организме, часто обусловлены не уменьшением концентрации вещества, а увеличением расстояния между средами, в частности, за счет утолщения тех или иных тканевых структур. В мертвых тканях градиенты падают. Только живое в состоянии поддерживать неравновесное состояние своих сред. Многие патологические процессы, связанные с нарушением транспорта веществ в организме, часто обусловлены не уменьшением концентрации вещества, а увеличением расстояния между средами, в частности, за счет утолщения тех или иных тканевых структур. В мертвых тканях градиенты падают. Только живое в состоянии поддерживать неравновесное состояние своих сред.

Концентрационные градиенты предопределяют стремление веществ перейти туда, где их концентрация ниже, Концентрационные градиенты предопределяют стремление веществ перейти туда, где их концентрация ниже, а осмотические заставляют воду переходить через мембрану в направлении более высокого осмотического давления.

Однако, наличие градиентов само по себе не означает, что вещества транспортируются через мембрану. Это в конечном счете определяется способностью мембраны пропускать через себя то или иное вещество. Разные мембраны неодинаково относятся к одним и тем же веществам. Эта способность мембран избирательно пропускать вещества обозначается термином - проницаемость мембран . Однако, наличие градиентов само по себе не означает, что вещества транспортируются через мембрану. Это в конечном счете определяется способностью мембраны пропускать через себя то или иное вещество. Разные мембраны неодинаково относятся к одним и тем же веществам. Эта способность мембран избирательно пропускать вещества обозначается термином - проницаемость мембран .

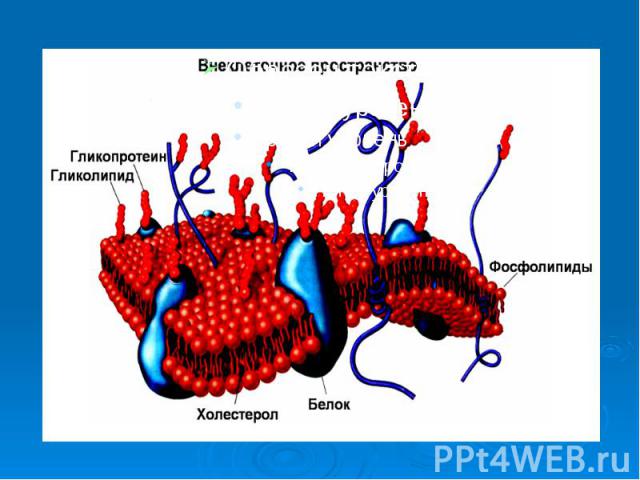
3. Ультраструктура и функции биологических мембран Проникновение веществ в клетку и из клетки в большой степени зависит от свойств мембраны. Мембраны образуют оболочки всех органелл клетки: ядра, митохондрий, лизосом, аппарата Гольджи, эндоплазматического ретикулума. Раньше считалось, что мембраны состоят только из тонкого слоя липидов. В экспериментах с эритроцитами установили, что площадь липидов, входящих в состав мембраны, вдвое больше площади самого эритроцита, в результате был сделан вывод, что мембраны состоят из двойного слоя фосфолипидов. (3)
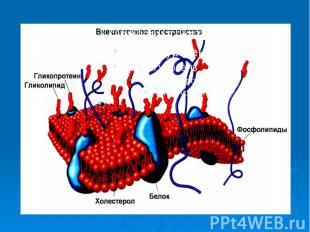


Такая молекула называется лизолицетин. Оказавшись в составе мембран, такие молекулы образуют пору, например, на рис. 2 показана пора 6. Мембрана теряет барьерные свойства. Это приводит к повышению проницаемости мембраны клеток для ионов натрия и некоторых других или к полному разрушению клеток. Например, при укусах некоторых змей наблюдается гемолиз эритроцитов. Проницаемости для всех ионов при образовании пор в мембране выравнивается. Потенциал покоя снижается . Митохондрия не может участвовать в синтезе АТФ, нервные клетки не могут проводить нервный импульс. Такая молекула называется лизолицетин. Оказавшись в составе мембран, такие молекулы образуют пору, например, на рис. 2 показана пора 6. Мембрана теряет барьерные свойства. Это приводит к повышению проницаемости мембраны клеток для ионов натрия и некоторых других или к полному разрушению клеток. Например, при укусах некоторых змей наблюдается гемолиз эритроцитов. Проницаемости для всех ионов при образовании пор в мембране выравнивается. Потенциал покоя снижается . Митохондрия не может участвовать в синтезе АТФ, нервные клетки не могут проводить нервный импульс. Подобные процессы происходят при вирусных и бактериальных инфекциях, действии ионизирующего излучения и т.д.

Действие многих лекарственных веществ направлено на восстановление барьерных свойств мембран клеток. Действие многих лекарственных веществ направлено на восстановление барьерных свойств мембран клеток. В заключении этого раздела перечислим основные функции биологических мембран: барьерная, матричная, механическая и специфические - генерация и проведение биопотенциалов действия, рецепторная, энергетическая

4. Пассивный транспорт

Перемещение веществ в клетку или из нее во внеклеточную среду может осуществляться многими способами. В зависимости от того, что является движущей силой перемещения, все виды перемещения веществ могут быть разделены на пассивный и активный транспорт. Пассивный транспорт - всегда за счет энергии, сконцентрированной в каком-нибудь градиенте, а не за счет АТФ. Пассивный всегда по направлению градиента, то есть от более высокого уровня к низкому Перемещение веществ в клетку или из нее во внеклеточную среду может осуществляться многими способами. В зависимости от того, что является движущей силой перемещения, все виды перемещения веществ могут быть разделены на пассивный и активный транспорт. Пассивный транспорт - всегда за счет энергии, сконцентрированной в каком-нибудь градиенте, а не за счет АТФ. Пассивный всегда по направлению градиента, то есть от более высокого уровня к низкому

Часто присутствуют несколько градиентов, тогда перенос вещества осуществляется по результирующей всех градиентов. Основные градиенты, присущие живым организмам - концентрационный, осмотический, электрический, гидростатический. В соответствии с этим имеются следующие виды пассивного транспорта: диффузия, осмос, электроосмос, аномальный осмос и фильтрация. Часто присутствуют несколько градиентов, тогда перенос вещества осуществляется по результирующей всех градиентов. Основные градиенты, присущие живым организмам - концентрационный, осмотический, электрический, гидростатический. В соответствии с этим имеются следующие виды пассивного транспорта: диффузия, осмос, электроосмос, аномальный осмос и фильтрация.

4.1. Диффузия

Основной механизм пассивного транспорта, обусловленный концентрационным градиентом - диффузия. Основной механизм пассивного транспорта, обусловленный концентрационным градиентом - диффузия. Диффузия - это самопроизвольный процесс проникновения веществ из области большей в область меньшей концентрации в результате теплового хаотического движения молекул.

Математическое описание дал Фик: скорость диффузии прямо пропорциональна градиенту концентрации и площади , через которую осуществляется диффузия.

Проникновение растворенных веществ, обладающих электрическим зарядом, зависит не только от концентрационного градиента, но и от электрического градиента мембраны и может возникнуть движение против концентрационного по электрическому градиенту. Совокупность концентрационного и электрического градиента называется электрохимическим градиентом Проникновение растворенных веществ, обладающих электрическим зарядом, зависит не только от концентрационного градиента, но и от электрического градиента мембраны и может возникнуть движение против концентрационного по электрическому градиенту. Совокупность концентрационного и электрического градиента называется электрохимическим градиентом

Помимо простой существует облегченная диффузия и ее разновидность – обменная диффузия. Рассмотрим несколько видов этого процесса: Помимо простой существует облегченная диффузия и ее разновидность – обменная диффузия. Рассмотрим несколько видов этого процесса:

4.1.1.Транспорт ионов с помощью подвижных переносчиков

Таким образом, например, переносит ионы калия антибиотик валиномицин. Молекула валиномицина, имеющая внутри полость, захватывает в нее ион калия на одной стороне мембраны и переносит этот ион на другую сторону мембраны. Таким образом, например, переносит ионы калия антибиотик валиномицин. Молекула валиномицина, имеющая внутри полость, захватывает в нее ион калия на одной стороне мембраны и переносит этот ион на другую сторону мембраны. В присутствии валиномицина отношение коэффициентов проницаемостей для ионов Na+ и К+ биологической мембраны равен:

4.1.2.Эстафетный перенос Эстафетный перенос при обменной диффузии осуществляется с помощью двух или нескольких носителей через мембрану частицы. При этом частица последовательно передается в мембране от одного носителя к другому.

Примером эстафетного переноса может служить движение через мембрану ионов натрия Na+ с помощью антибиотика грамицидина.Грамицидин, создающий в мембране полуканал,

Диффундируя вдоль мембраны, молекула грамицидина встречается с другой молекулой грамицидина, находящейся на внутренней стороне мембраны, и передает ей ион натрия. Молекулы грамицидина образуют временную цепочку поперек мембраны и «по эстафете» передают ионы Na+ от одной молекулы переносчика к другой. Затем ион натрия выбрасывается внутрь клетки. Так как грамицидин фактически увеличивает проницаемость мембраны для ионов Na+, т.е. нарушает нормальное функционирование клетки, он используется в виде мази как контрацептив, блокирующий функционирование сперматозоидов и яйцеклеток. Вещества, облегчающие перенос ионов через мембрану, называются ионофорами (нонактин, валиномицин, грамицидин, моноксин и т.д.). Диффундируя вдоль мембраны, молекула грамицидина встречается с другой молекулой грамицидина, находящейся на внутренней стороне мембраны, и передает ей ион натрия. Молекулы грамицидина образуют временную цепочку поперек мембраны и «по эстафете» передают ионы Na+ от одной молекулы переносчика к другой. Затем ион натрия выбрасывается внутрь клетки. Так как грамицидин фактически увеличивает проницаемость мембраны для ионов Na+, т.е. нарушает нормальное функционирование клетки, он используется в виде мази как контрацептив, блокирующий функционирование сперматозоидов и яйцеклеток. Вещества, облегчающие перенос ионов через мембрану, называются ионофорами (нонактин, валиномицин, грамицидин, моноксин и т.д.).

Одна из главных особенностей пассивного транспорта - его избирательность. Существуют переносчики для D-глюкозы, но нет переносчиков для L-глюкозы. В случае простой диффузии избирательность определяется сродством переносимой частицы к гидрофильной голове фосфолипида. Одна из главных особенностей пассивного транспорта - его избирательность. Существуют переносчики для D-глюкозы, но нет переносчиков для L-глюкозы. В случае простой диффузии избирательность определяется сродством переносимой частицы к гидрофильной голове фосфолипида.

4.1.3. Перенос ионов через каналы биомембран
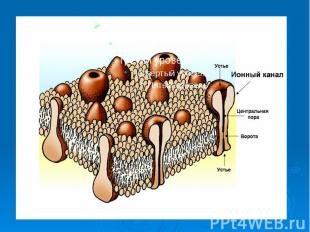

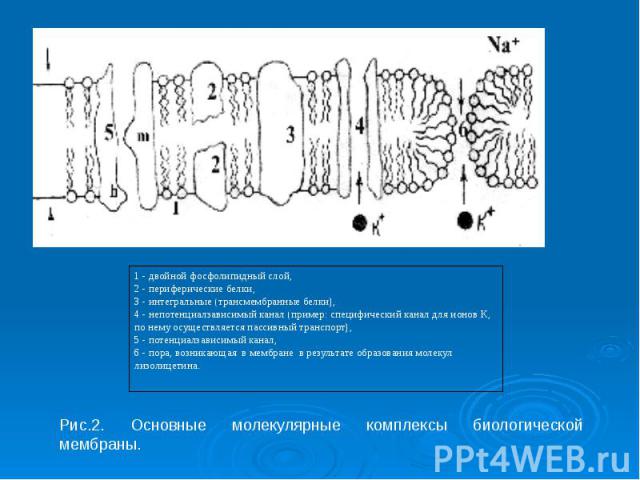
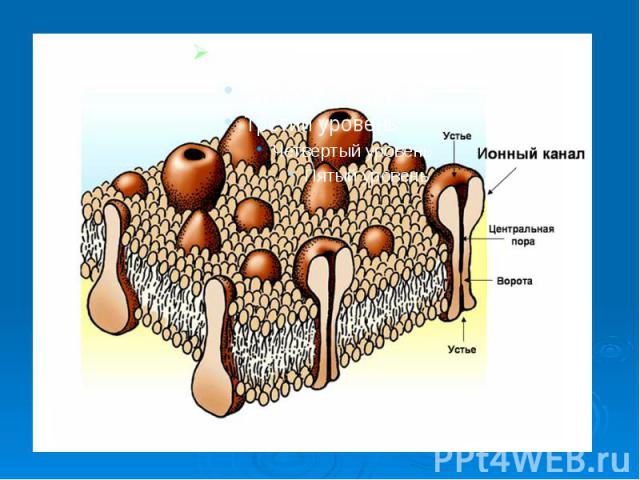
В основе многих физиологических процессов (передача электрических и химических сигналов, мышечное сокращение, секреторный процесс и т.д. ) лежит прежде всего работа ионных каналов. Их В основе многих физиологических процессов (передача электрических и химических сигналов, мышечное сокращение, секреторный процесс и т.д. ) лежит прежде всего работа ионных каналов. Их характеристики могут изменять некоторые фармакологические препараты и яды. Существуют блокаторы ионных каналов, например, лекарственные вещества, антагонисты кальция (верапамил, нифедипин и др.) они временно блокируют ионные каналы, снижая мышечный тонус сосудов. Есть лекарственные вещества, временно блокирующие натриевые каналы, например, анестетик - лидокаин, новокаин. Они снижают местную чувствительность, устраняют чувство боли.

Необратимо блокируют Na - ионные каналы такие паралитические яды, как тетродотоксин, вырабатываемый рыбой Spheroidus rubrides (фугу) или сакситоксин, продуцируемый некоторыми планктонами. Необратимо блокируют Na - ионные каналы такие паралитические яды, как тетродотоксин, вырабатываемый рыбой Spheroidus rubrides (фугу) или сакситоксин, продуцируемый некоторыми планктонами.

Сильный блокирующий эффект калиевых ионных каналов оказывает тетраэтиламмоний. Сильный блокирующий эффект калиевых ионных каналов оказывает тетраэтиламмоний. Существуют также активаторы, открывающие ионные каналы, например, лекарственный препарат миноксидил, активирует калиевые каналы.

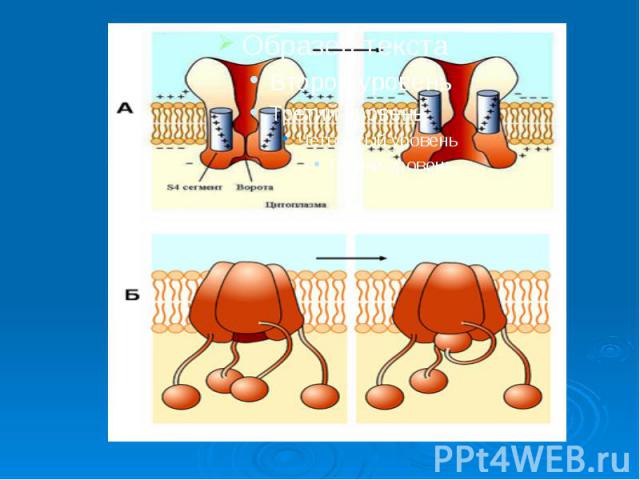
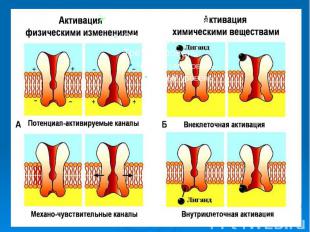
Каналам биомембран свойственна характерная избирательность для ионов (селективность), а также способность открываться и закрываться при различных воздействиях на мембрану (воротная функция). Переходы каналов из открытого состояния в закрытое (воротный механизм) могут быть обусловлены изменениями мембранного потенциала, взаимодействием с определенными химическими веществами, специфическим фосфорилированием каналов Каналам биомембран свойственна характерная избирательность для ионов (селективность), а также способность открываться и закрываться при различных воздействиях на мембрану (воротная функция). Переходы каналов из открытого состояния в закрытое (воротный механизм) могут быть обусловлены изменениями мембранного потенциала, взаимодействием с определенными химическими веществами, специфическим фосфорилированием каналов

Кроме вышеописанных каналов в мембране находятся неспецифические каналы для пассивного транспорта ионов (в первую очередь для ионов калия). Эти каналы не имеют воротных механизмов, всегда открыты и почти не меняют свое состояние при электрических воздействиях на мембрану. Кроме вышеописанных каналов в мембране находятся неспецифические каналы для пассивного транспорта ионов (в первую очередь для ионов калия). Эти каналы не имеют воротных механизмов, всегда открыты и почти не меняют свое состояние при электрических воздействиях на мембрану.

4.2.Осмос Клеточные мембраны обладают свойством полупроницаемости, то есть способностью хорошо пропускать одни вещества, например, воду, и плохо другие. Вода проходит в результате осмоса

Осмос- это движение молекулы воды через полупроницаемую мембрану из области меньшей в область большей концентрации растворенного вещества. Сила вызывающая это движение - осмотическое давление. Осмос- это движение молекулы воды через полупроницаемую мембрану из области меньшей в область большей концентрации растворенного вещества. Сила вызывающая это движение - осмотическое давление.

Осмос, по существу, представляет собой диффузию молекул растворителя. За меру осмотического давления принимают то механическое давле­ние, например гидростатическое, которое уравновешивает осмоти­ческое давление и выравнивает потоки молекул растворителя в одну и другую стороны. Осмотическое давление зависит от количества растворенных частиц и от температуры. Осмос, по существу, представляет собой диффузию молекул растворителя. За меру осмотического давления принимают то механическое давле­ние, например гидростатическое, которое уравновешивает осмоти­ческое давление и выравнивает потоки молекул растворителя в одну и другую стороны. Осмотическое давление зависит от количества растворенных частиц и от температуры.

P=iRCT R- универсальная газовая постоянная, P=iRCT R- универсальная газовая постоянная, i - изотонический коэффициент - показывает во сколько раз увеличи­вается количество частиц при диссоциации молекулы вещества. Для неэлектролитов i=1. Осм. давление крови человека 7,6–7,8 атм.

4.3.Фильтрация Кроме осмоса, перенос воды может осуществляться путем фильтрации при наличии градиента гидростатического давления. Фильтрация - движение жидкости через поры какой-либо перего­родки под действием гидростатического давления.

Фильтрация - движение жидкости через поры какой-либо перего­родки под действием гидростатического давления. под давлением. Фильтрация - движение жидкости через поры какой-либо перего­родки под действием гидростатического давления. под давлением.

4.4.Водный обмен между кровью и лимфой Явление фильтрации и осмоса имеют особое значение в процессе об­мена воды между кровью и тканями. Осмотическое давление крови человека 7,6 - 7,8 атмосфер. Это давление является суммой давлений всех растворенных в плазме крови веществ.

Особое значение в водном обмене между кровью и тканевой жидкостью имеет та часть общего осмотического давления, которая обусловлена высокомоле­кулярными веществами - белками. Эта часть осмотического давле­ния называется онкотическим давлением. Величина осмотического давления в 200 раз превосходит величину онкотического давления. Несмотря на это онкотическому давлению принадлежит основная роль в поступлении воды в кровеносное русло из тканевой жид Особое значение в водном обмене между кровью и тканевой жидкостью имеет та часть общего осмотического давления, которая обусловлена высокомоле­кулярными веществами - белками. Эта часть осмотического давле­ния называется онкотическим давлением. Величина осмотического давления в 200 раз превосходит величину онкотического давления. Несмотря на это онкотическому давлению принадлежит основная роль в поступлении воды в кровеносное русло из тканевой жид

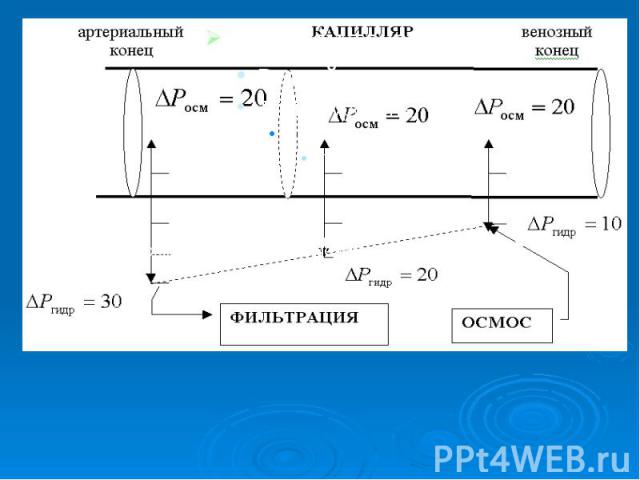
Онкотичесое давление крови человека равно 30 мм.рт.ст. ,а тканевой жид­кости и лимфы 10 мм.рт.ст. Под действием разности онкотического давления крови и лимфы величиной 20 мм.рт.ст. вода поступает из лимфы в кровь. Одновременно с градиентом онкотического давления () существует и градиент гидростатического давления, обусловленный работой сердца (). В артериальном конце капилляра гидростатическое давление крови 30 мм.рт.ст.. На протяжении капилляра кровяное давление падает: в середине оно равно 20 мм.рт.ст, а в венозном конце - 10 мм.рт.ст.. Онкотичесое давление крови человека равно 30 мм.рт.ст. ,а тканевой жид­кости и лимфы 10 мм.рт.ст. Под действием разности онкотического давления крови и лимфы величиной 20 мм.рт.ст. вода поступает из лимфы в кровь. Одновременно с градиентом онкотического давления () существует и градиент гидростатического давления, обусловленный работой сердца (). В артериальном конце капилляра гидростатическое давление крови 30 мм.рт.ст.. На протяжении капилляра кровяное давление падает: в середине оно равно 20 мм.рт.ст, а в венозном конце - 10 мм.рт.ст..
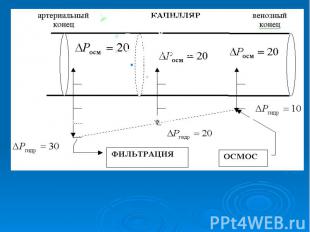

Соответственно, если сравнить значения градиентов онкотического и гидростатического давлений в различных участках капилляров, то очевидно, что в артериальном конце преобладает гидростатическое давление - наблюдается фильтрация, в результате которой вода выходит из кровяного русла, а в венозном конце преобладает онкотическое давление, т.е. наблюдается осмос, и вода поступает из ткани в лимфу Соответственно, если сравнить значения градиентов онкотического и гидростатического давлений в различных участках капилляров, то очевидно, что в артериальном конце преобладает гидростатическое давление - наблюдается фильтрация, в результате которой вода выходит из кровяного русла, а в венозном конце преобладает онкотическое давление, т.е. наблюдается осмос, и вода поступает из ткани в лимфу

В норме процессы фильтрации и осмоса скомпенсированы. При патологиях, например: лучевая болезнь – нарушение проницаемости мембран, при долгих голоданиях, гипертонии, шоке, ожогах, при больших кровопотерях уменьшается градиент онкотического давления, и тогда фильтрация воды преобладает над осмосом, вследствие этого наблюдаются отеки. В норме процессы фильтрации и осмоса скомпенсированы. При патологиях, например: лучевая болезнь – нарушение проницаемости мембран, при долгих голоданиях, гипертонии, шоке, ожогах, при больших кровопотерях уменьшается градиент онкотического давления, и тогда фильтрация воды преобладает над осмосом, вследствие этого наблюдаются отеки.

5. Активный транспорт Перенос молекул и ионов против электрохимического градиента, осуществляемый клеткой за счет энергии метаболических процессов, называется активным транспортом. Осуществляя такой транспорт, клетка совершает работу, которая называется концентрационной или осмотической. Накопление клеткой вещества, содержащегося в окружающей среде в малых количествах, и выведение тех веществ, концентрация которых в окружающей среде значительно больше, чем в клетке, обеспечивается активным транспортом.

A=mRTlnС1/C2 A-концентрационная работа, m-количество молей вещества, перенесенных через мембрану, T -абсолютная температура, -С1 и С2 концентрации ионов внутри и вне клетки.

У человека в покое примерно 30 - 40 % всей энергии, образующейся в ходе метаболических процессов, расходуется на активный транспорт. В тканях, где активный транспорт особенно интенсивен, потребляется много кислорода даже в покое; так мозг человека составляет 1/50 массы тела, а потребляет в покое 1/5 всего кислорода. Активный транспорт - одно из удивительных свойств живых организмов. У человека в покое примерно 30 - 40 % всей энергии, образующейся в ходе метаболических процессов, расходуется на активный транспорт. В тканях, где активный транспорт особенно интенсивен, потребляется много кислорода даже в покое; так мозг человека составляет 1/50 массы тела, а потребляет в покое 1/5 всего кислорода. Активный транспорт - одно из удивительных свойств живых организмов.

Активный транспорт веществ через биологические мембраны имеет огромное значение. За счет активного транспорта в организме создаются градиенты концентраций, градиенты электрических потенциалов, градиенты давления и т.д., поддерживающие жизненные процессы, то есть с точки зрения термодинамики активный перенос удерживает организм в неравновесном состоянии, поддерживает жизнь. Активный транспорт веществ через биологические мембраны имеет огромное значение. За счет активного транспорта в организме создаются градиенты концентраций, градиенты электрических потенциалов, градиенты давления и т.д., поддерживающие жизненные процессы, то есть с точки зрения термодинамики активный перенос удерживает организм в неравновесном состоянии, поддерживает жизнь.

Изучение систем активного транспорта, называемых биологическими насосами, является важной задачей исследования клеточных мембран. Нарушение активного транспорта приводит к гибели клетки. Отключение Nа+- К+ насоса на 10 минут приводит к изменению ионного состава внутри клетки на 40 - 50 %. Изучение систем активного транспорта, называемых биологическими насосами, является важной задачей исследования клеточных мембран. Нарушение активного транспорта приводит к гибели клетки. Отключение Nа+- К+ насоса на 10 минут приводит к изменению ионного состава внутри клетки на 40 - 50 %.

В покое мембрана не является абсолютно непроницаемой для любого вида ионов. Диффузионные потоки, не будь им противодей­ствия, рано или поздно выровняли бы состав по обе стороны мем­браны. Для восстановления и поддержания необходимой концентра­ции ионов внутри клетки в мембране содержится специальный аппа­рат, обеспечивающий их активный транспорт против градиента их собственной концентрации. Активный транспорт ионов Nа+, К+ и Са+ осуществляется двумя типами "насосов" Nа+ - К+ и Nа+ - Са2+. В покое мембрана не является абсолютно непроницаемой для любого вида ионов. Диффузионные потоки, не будь им противодей­ствия, рано или поздно выровняли бы состав по обе стороны мем­браны. Для восстановления и поддержания необходимой концентра­ции ионов внутри клетки в мембране содержится специальный аппа­рат, обеспечивающий их активный транспорт против градиента их собственной концентрации. Активный транспорт ионов Nа+, К+ и Са+ осуществляется двумя типами "насосов" Nа+ - К+ и Nа+ - Са2+.

Наиболее характерная черта таких насосов - перенос вещества против градиентов. Следовательно, необходимо энергетическое обеспече­ние такого процесса. Организм должен за счет своей свободной энергии преодолеть физико-химические градиенты, повернуть вспять движение веществ. Значит, второй особенностью активного транс­порта является наличие источника энергии для его осуществления. Причем эта энергия поступает из химических реакций Наиболее характерная черта таких насосов - перенос вещества против градиентов. Следовательно, необходимо энергетическое обеспече­ние такого процесса. Организм должен за счет своей свободной энергии преодолеть физико-химические градиенты, повернуть вспять движение веществ. Значит, второй особенностью активного транс­порта является наличие источника энергии для его осуществления. Причем эта энергия поступает из химических реакций

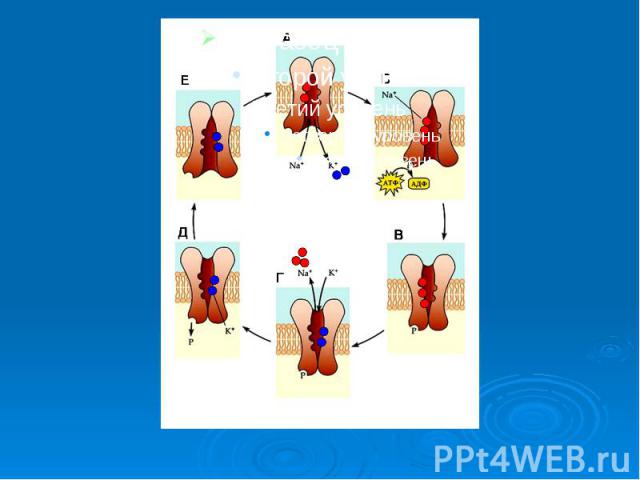
Рассмотрим этот процесс на примере К+-Nа насоса. Ионы К+ и Nа+ вводятся и выводятся из клетки и в клетку против концен­трационного градиента. Клетка очень строго следит за содержанием в своей цитоплазме названных ионов Рассмотрим этот процесс на примере К+-Nа насоса. Ионы К+ и Nа+ вводятся и выводятся из клетки и в клетку против концен­трационного градиента. Клетка очень строго следит за содержанием в своей цитоплазме названных ионов
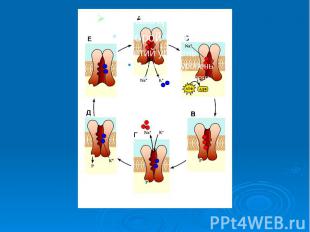

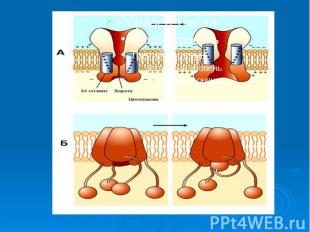
При повышении концентрации ионов натрия активируется так называемая калий-натрий зависимая АТФ-аза, этот фермент содержащийся в плазматической мембране, активирует гидролиз АТФ. При повышении концентрации ионов натрия активируется так называемая калий-натрий зависимая АТФ-аза, этот фермент содержащийся в плазматической мембране, активирует гидролиз АТФ. Активируя гидролиз АТФ, он обеспечивает фосфорилирование переносчика, который выносит натрий из клетки. На внешней поверхности мембраны происходит ионный обмен натрия на калий внутри ферментного комплекса, после этого обмена белок-переносчик совершает обратный переворот с переносом калия внутрь клетки. Ион калия и неорганическая фосфатная группа (Р) освобождается от белка-переносчика, который в свою очередь возвращается в исходное состояние. Подсчитано, что за полный цикл 1 молекула АТФ обеспечи­вает перенос через клеточную мембрану 3 иона Na+ и 2 иона К+.

Факторы, характеризующие систему активного транспорта. 1. Перенос веществ против электро-химических градиентов. 2. Наличие источника энергии (АТФ). 3. Необходимость переносчика вещества (молекула - белковой природы). Присутствие фермента, который активирует этот процесс – транспортной АТФ-азы. Однако вся эта система не работает до появления определенного стимула, которым обычно служит нарастающая концентрация транспортируемого вещества.

Роль активного транспорта в физиологическом процессе Активный транспорт необходим: Для поддержания высокой концентрации ионов K+ внутри клетки, этим самым он обеспечивает постоянство величины потенциала покоя (см. раздел 2.3.). Для поддержания низкой концентрации ионов Na+ внутри клетки, что, с одной стороны, обеспечивает работу механизма генерации потенциала действия, а с другой стороны, обеспечивает сохранение нормальной осмолярности объема клетки (см. раздел 2.4.). Поддерживая стабильный концентрационный градиент Na+, Na+ – K+ насос способствует напряженному транспорту аминокислот, сахаров через клеточную мембрану.

БИОФИЗИКА КЛЕТКИ ЛЕКЦИЯ 2 Биоэлектрические потенциалы

План лекции 1.Методы измерения потенциала 2.Модель Нернста 3. Потенциал покоя 4. Потенциал действия 5. Распространение нервного импульса вдоль возбудимого волокна

1. Методы измерения мембранного потенциала. Это явление лежит в основе возбудимости клеток, регуляции внутриклеточных процессов, работы нервной Одна из важнейших функций биологической мембраны - генерация и передача биопотенциалов. системы, регуляции мышечного сокращения, рецепции. В медицине на исследовании электрических полей, созданных биопотенциалами органов и тканей, основаны диагностические методы: электрокардиография, электроэнцефалография, электромиография и другие. Практикуется и лечебное воздействие на ткани и органы внешними электрическими импульсами при электростимуляции.


В процессе жизнедеятельности в клетках и тканях могут возникать разности электрических потенциалов: В процессе жизнедеятельности в клетках и тканях могут возникать разности электрических потенциалов: окислительно-восстановительные потенциалы - вследствие переноса электронов от одних молекул к другим; мембранные - вследствие градиента концентрации ионов и переноса ионов через мембрану. Биопотенциалы, регистрируемые в организме, - это в основном мембранные потенциалы.

Мембранным потенциалом называется разность потенциалов между внутренней (цитоплазматической) и наружной поверхностями мембраны: Мембранным потенциалом называется разность потенциалов между внутренней (цитоплазматической) и наружной поверхностями мембраны: . В дальнейшем для упрощения написания формул величину будем обозначать просто как .

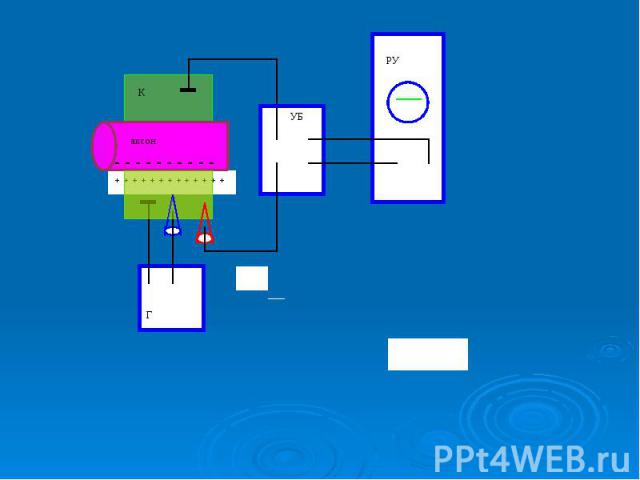
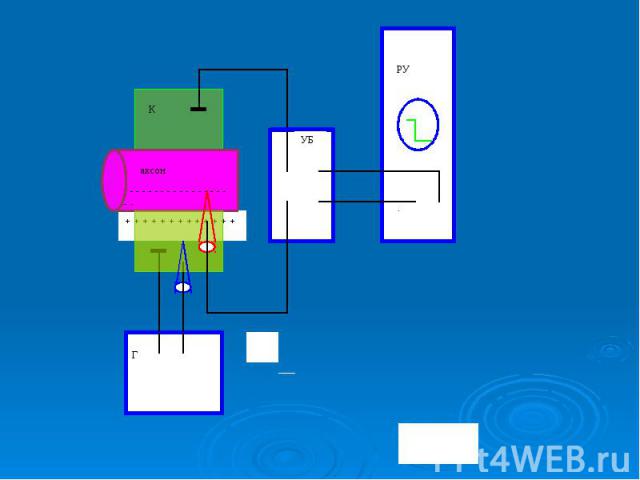
Прогресс в исследовании биопотенциалов обусловлен: Прогресс в исследовании биопотенциалов обусловлен: разработкой микроэлектродного метода внутриклеточного измерения потенциалов; созданием специальных усилителей биопотенциалов (УПТ); выбором удачных объектов исследования крупных клеток и среди них гигантского аксона кальмара. Диаметр аксона кальмара достигает 0,5 мм, что в 100-1000 больше, чем диаметр аксонов позвоночных животных, в том числе человека. Гигантские, в сравнении с позвоночными, размеры аксона этого проворного и ловкого головоногого моллюска имеют большое физиологическое значение - обеспечивают быструю передачу нервного импульса по нервному волокну. Для биофизики гигантский аксон кальмара послужил великолепным модельным объектом для изучения биопотенциалов (недаром выдвигались предложения поставить памятник кальмару - животному, которому так многим обязана наука, подобно существующим памятникам лягушке в Париже и собаке под Санкт-Петербургом).

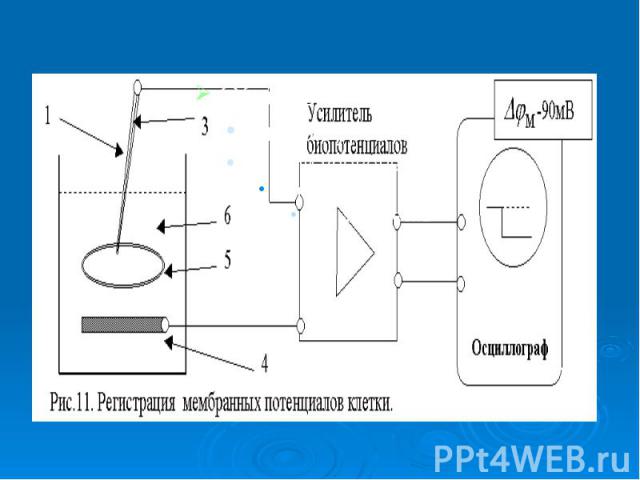
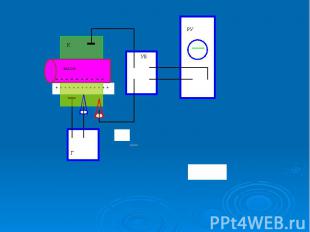
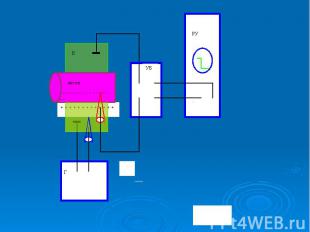
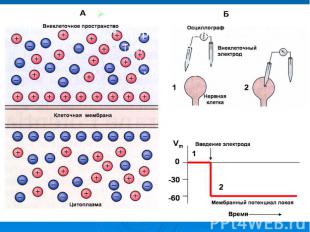
В гигантский аксон кальмара можно ввести микроэлектрод, не нанеся аксону значительных повреждений. В гигантский аксон кальмара можно ввести микроэлектрод, не нанеся аксону значительных повреждений. Стеклянный микроэлектрод (1) представляет собой стеклянную микропипетку с оттянутым очень тонким кончиком (диаметр 0,1-0,5 мкм).

Металлический электрод такой толщины пластичен, и не может проколоть клеточную мембрану, кроме того, он поляризуется. Для исключения поляризации электрода используются неполяризующиеся электроды, например, серебряная проволока (3), покрытая солью , помещенные в раствор или , заполняющий микроэлектрод Металлический электрод такой толщины пластичен, и не может проколоть клеточную мембрану, кроме того, он поляризуется. Для исключения поляризации электрода используются неполяризующиеся электроды, например, серебряная проволока (3), покрытая солью , помещенные в раствор или , заполняющий микроэлектрод
 - располагается в растворе (6) у наружной поверхно")
Второй электрод сравнения (4) - располагается в растворе (6) у наружной поверхности клетки (5) (рис. 11). Регистрирующее устройство осциллограф. Второй электрод сравнения (4) - располагается в растворе (6) у наружной поверхности клетки (5) (рис. 11). Регистрирующее устройство осциллограф.
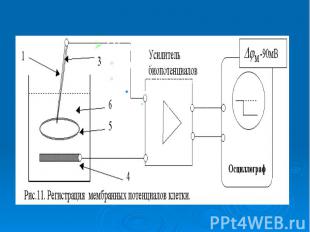

Микроэлектродный метод дал возможность измерить биопотенциалы не только на гигантском аксоне кальмара, но и на клетках нормальных размеров: нервных волокнах других животных, клетках скелетных мышц, клетках миокарда и других Микроэлектродный метод дал возможность измерить биопотенциалы не только на гигантском аксоне кальмара, но и на клетках нормальных размеров: нервных волокнах других животных, клетках скелетных мышц, клетках миокарда и других
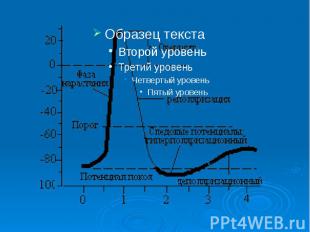

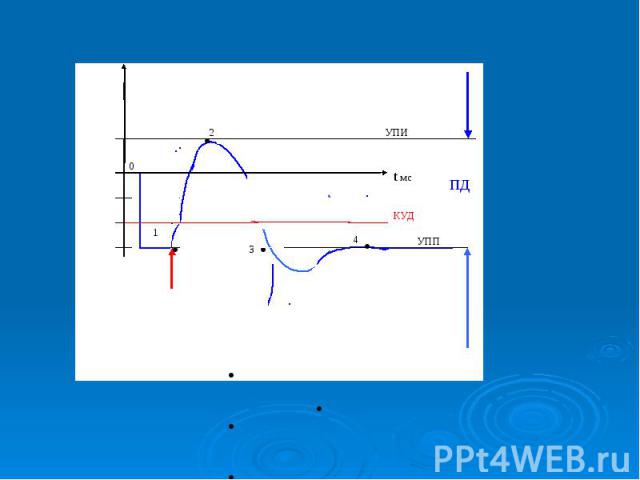
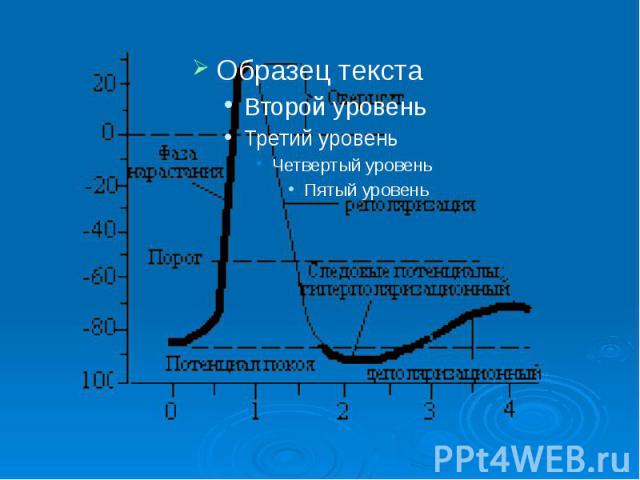
Биопотенциалы, электрическая активность клеток животного и растительного происхождения, обусловленная неравномерным распределением электролитов внутри и вне клеток. Мембранный потенциал (МП, потенциал покоя) определяется трансмембранным градиентом концентрации калия (К) (внутри клеток концентрация К выше) и остается постоянным длительное время, пока клетка не активируется внешним воздействием. При этом внутренняя часть клеток имеет отрицательный заряд. Переход клеток в активное состояние вызывается быстрым сдвигом МП в положительном направлении - потенциалом действия (ПД), для которого характерно несколько фаз: фаза деполяризации, обусловленная входом натрия (Na) внутрь клеток, вызывающим изменение поляризации клетки, - овершут; фаза реполяризации, в течение которой восстанавливается исходный потенциал вследствие выхода К из клеток; следовые (деполяризационные и гиперполяризационные) потенциалы. Постоянство и восстановление исходного распределения электролитов внутри клеток обеспечивается Na/K насосом. На рис. - временный ход потенциала действия.

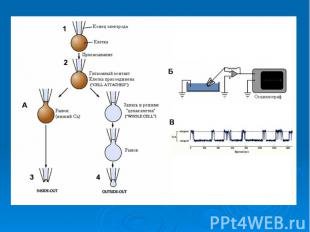
Другим эффективным методом измерения потенциала мембраны стал метод локальной фиксации потенциала ("Patch Clamp"). Суть метода заключается в том, что микроэлектрод тонким концом, имеющим диаметр 0,5-1 мкм, присасывается к мембране таким образом, чтобы в его внутренний диаметр попал ионный канал. Тогда, используя схему фиксации потенциала, можно измерять токи, которые проходят только через одиночный канал мембраны, а не через все каналы одновременно, как это происходит при использовании стандартного метода фиксации потенциала, описанного выше. Другим эффективным методом измерения потенциала мембраны стал метод локальной фиксации потенциала ("Patch Clamp"). Суть метода заключается в том, что микроэлектрод тонким концом, имеющим диаметр 0,5-1 мкм, присасывается к мембране таким образом, чтобы в его внутренний диаметр попал ионный канал. Тогда, используя схему фиксации потенциала, можно измерять токи, которые проходят только через одиночный канал мембраны, а не через все каналы одновременно, как это происходит при использовании стандартного метода фиксации потенциала, описанного выше.

2. Элемент Нернста Самой простой и адекватной моделью является элемент Нернста (Рис.12.). Сосуд, в котором находится растворы одной и той же соли, но разной концентрации, разделен пористой перегородкой (C1 > C2).
. Вследствие разницы концентрации, ионы диффундируют, причем один из ионов, как")
). Вследствие разницы концентрации, ионы диффундируют, причем один из ионов, как правило, катион диффундирует быстрее и более разбавленный раствор приобретает знак катиона. По обе стороны перегородки образуется разность потенциалов, которую называют диффузионной разностью потенциалов. ). Вследствие разницы концентрации, ионы диффундируют, причем один из ионов, как правило, катион диффундирует быстрее и более разбавленный раствор приобретает знак катиона. По обе стороны перегородки образуется разность потенциалов, которую называют диффузионной разностью потенциалов.

При этом ЭДС, возникающая в концентрационном элементе, образованном раствором одной соли, определяется из уравнения Нернста. При этом ЭДС, возникающая в концентрационном элементе, образованном раствором одной соли, определяется из уравнения Нернста. - газовая постоянная (R=8,316 Дж/к-моль), - абсолютная температура, - число Фарадея (96500 кл/моль), n- валентность, С1 и С2 молекулярные концентрации ионов по обе стороны мембраны.

Уравнение Нернста

В реальных условиях, в клетке, разница скорости движения анионов и катионов обусловлена присутствием полупроницаемой мембраны. Без нее ЭДС концентрированного элемента быстро падает. Поэтому потенциалы, возникающие в таком элементе, называются не диффузионными, а мембранными. В реальных условиях, в клетке, разница скорости движения анионов и катионов обусловлена присутствием полупроницаемой мембраны. Без нее ЭДС концентрированного элемента быстро падает. Поэтому потенциалы, возникающие в таком элементе, называются не диффузионными, а мембранными.

Элемент Нернста

Теория электролитической диссоциации была впервые применена и объяснена в механизме биоэлектрогенеза В.Ю.Чаговцем (1896) слушателям ВМА, в дальнейшем развил учение Ю.Бернштейн и лауреаты Нобелевской премии (Ходжкин, Кац, Хакси). Теория электролитической диссоциации была впервые применена и объяснена в механизме биоэлектрогенеза В.Ю.Чаговцем (1896) слушателям ВМА, в дальнейшем развил учение Ю.Бернштейн и лауреаты Нобелевской премии (Ходжкин, Кац, Хакси).

3. Потенциал покоя. Установлено, что цитоплазма клеток существенно отличается по химическому составу от внеклеточной жидкости, находящейся в межклеточном пространстве (рис. 13.)

Ионы Среда Ионы Среда Внеклеточная Внутриклеточная К 1 41 Na 10 1 Cl 13 1 Рис. 13. Относительная концентрация ионов внутри и вне клетки.
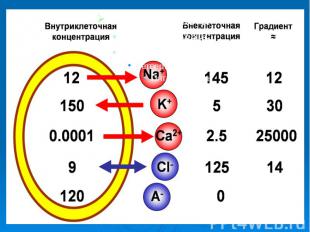

Разность ионных концентраций создает силу, стремящуюся выров­нять содержание ионов, но мембрана препятствует этому процессу. Мембраны обес­печивают избирательную проницаемость для различных ионов. Вследствие этого в тканях наблюдается неодинаковая скорость диффу­зии через клеточные мембраны катионов и анионов, что при нали­чии концентрационных градиентов служит непосредственной причиной возникновения мембранных потенциалов. Их называют биопотенциалами Разность ионных концентраций создает силу, стремящуюся выров­нять содержание ионов, но мембрана препятствует этому процессу. Мембраны обес­печивают избирательную проницаемость для различных ионов. Вследствие этого в тканях наблюдается неодинаковая скорость диффу­зии через клеточные мембраны катионов и анионов, что при нали­чии концентрационных градиентов служит непосредственной причиной возникновения мембранных потенциалов. Их называют биопотенциалами

Таким образом, источником электричества в живой ткани служит концентрационный элемент, образованный растворами электролита, имеющего неодинаковую концентрацию в цитоплазме и межклеточной жидкостью. При рассмотрении живых тканей в качестве концентра­ционного элемента необходимо учитывать диффузию через клеточ­ную мембрану не одного иона, а всех, концентрации которых нео­динаковы внутри и вне клетки. Это обстоятельство учитывает уравнение Гольдмана-Ходжкина-Катца. Таким образом, источником электричества в живой ткани служит концентрационный элемент, образованный растворами электролита, имеющего неодинаковую концентрацию в цитоплазме и межклеточной жидкостью. При рассмотрении живых тканей в качестве концентра­ционного элемента необходимо учитывать диффузию через клеточ­ную мембрану не одного иона, а всех, концентрации которых нео­динаковы внутри и вне клетки. Это обстоятельство учитывает уравнение Гольдмана-Ходжкина-Катца.

уравнение Гольдмана-Ходжкина-Катца.

Уравнение Гольдмана позволяет рассчитать разность потенциалов, существующую в разных клетках между цитоплазмой и межклеточной средой в покое и при возбуждении. Уравнение Гольдмана позволяет рассчитать разность потенциалов, существующую в разных клетках между цитоплазмой и межклеточной средой в покое и при возбуждении.

Потенциал покоя - стационарная разность электрических потенциалов, регистрируемая между внутренней и наружной поверхностями мембраны в невозбужденном состоянии Потенциал покоя - стационарная разность электрических потенциалов, регистрируемая между внутренней и наружной поверхностями мембраны в невозбужденном состоянии

Потенциал покоя определяется разной концентрацией ионов по разные стороны мембраны и диффузией ионов через мембрану. Результаты расчетов приблизительно совпадают с величинами мембранных потенциалов, полученными опытным путем Потенциал покоя определяется разной концентрацией ионов по разные стороны мембраны и диффузией ионов через мембрану. Результаты расчетов приблизительно совпадают с величинами мембранных потенциалов, полученными опытным путем

Проанализируем уравнение Гольдмана в состояние покоя: Проанализируем уравнение Гольдмана в состояние покоя: В состоянии покоя проницаемость мембраны для ионов значительно больше, чем для , и больше, чем для

Для аксона кальмара, например, Для аксона кальмара, например, Из уравнения Гольдмана получим уравнение Нернста для мембранного потенциала покоя: ;

Таким образом, уравнение Нернста - частный случай уравнения Гольдмана. При концентрации в клетке 20 мМ и аксоплазме 400 мМ рассчитанное значение равно - 95мВ. Действительно, в опытах на аксонах регистрируют потенциалы покоя около - 90мВ. Следовательно, в состоянии покоя мембранный потенциал (ПП) равен равновесному электрохимическому потенциалу по. Таким образом, уравнение Нернста - частный случай уравнения Гольдмана. При концентрации в клетке 20 мМ и аксоплазме 400 мМ рассчитанное значение равно - 95мВ. Действительно, в опытах на аксонах регистрируют потенциалы покоя около - 90мВ. Следовательно, в состоянии покоя мембранный потенциал (ПП) равен равновесному электрохимическому потенциалу по.

Итак, все клетки в состоянии покоя характеризуются определенной степенью поляризации. Клеточная мембрана всегда заряжена. ПП в различных клетках различен и достигает нескольких десятков милливольт. У аксона кальмара ПП-85мВ, у нервных и мышечных волокон ПП-90мВ. Итак, все клетки в состоянии покоя характеризуются определенной степенью поляризации. Клеточная мембрана всегда заряжена. ПП в различных клетках различен и достигает нескольких десятков милливольт. У аксона кальмара ПП-85мВ, у нервных и мышечных волокон ПП-90мВ.


4.Потенциал действия Все клетки возбудимых тканей при действии различных раздражителей достаточной силы переходят в состояние возбуждения. Возбудимость - это способность клеток к быстрому ответу на раздраже­ние, проявляющаяся через совокупность физических, физико-хими­ческих процессов и функциональных изменений

Обязательным признаком возбуждения является изменение электрического состо­яния клеточной мембраны. Опыт показывает, что возбужденный участок клетки становится электроотрицательным по отношению к невозбужденному, что указывает на перераспределение ионов в воз­бужденном участке. Оно имеет временный характер Обязательным признаком возбуждения является изменение электрического состо­яния клеточной мембраны. Опыт показывает, что возбужденный участок клетки становится электроотрицательным по отношению к невозбужденному, что указывает на перераспределение ионов в воз­бужденном участке. Оно имеет временный характер

Общее изменение разности потенциалов между клеткой и средой, происходящее при возбуждении, называется потенциалом действия Общее изменение разности потенциалов между клеткой и средой, происходящее при возбуждении, называется потенциалом действия
, или потенциал возбуждения нервных клеток (волокон), воз")
Потенциал действия (ПД), или потенциал возбуждения нервных клеток (волокон), возникает в ответ на достаточное по силе раздражение. ПД - очень быстрый, кратковременный электрический процесс, поэтому для его регистрации необходим катодный осциллограф с широкополосным усилителем. Потенциал действия (ПД), или потенциал возбуждения нервных клеток (волокон), возникает в ответ на достаточное по силе раздражение. ПД - очень быстрый, кратковременный электрический процесс, поэтому для его регистрации необходим катодный осциллограф с широкополосным усилителем.

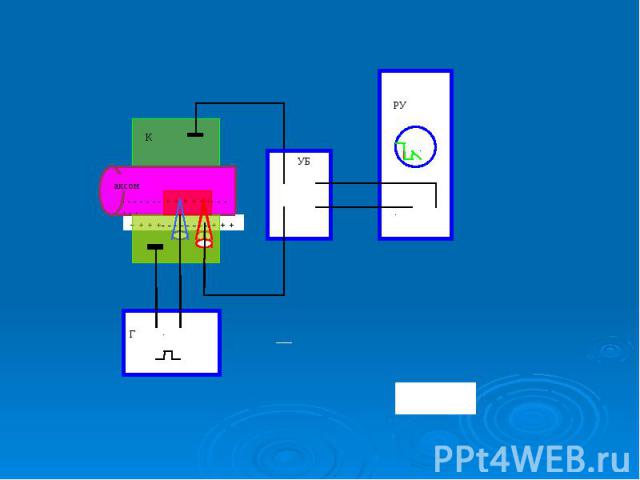
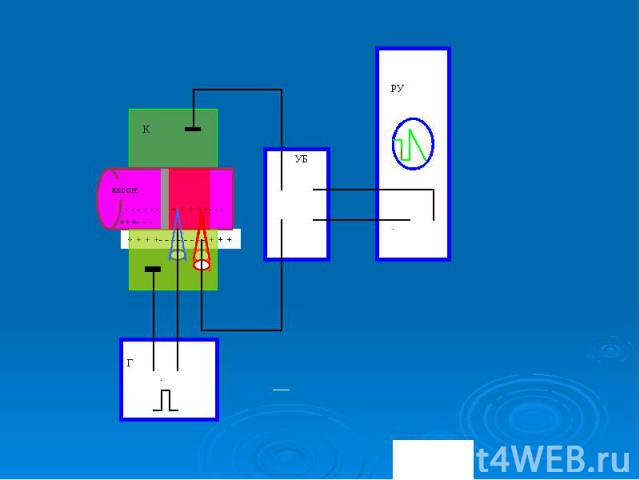
Классическое исследование параметров и механизма ПД проделано на гигантском аксоне кальмара с внутриклеточным раздражением и отведением внутриклеточного потенциала. В это нервное волокно (диаметром 0,5-1 мм) вводили на всю его длину два тончайших проволочных электрода. Один из них был раздражающим: через него в волокно подавали толчки тока того или иного направления, другой регистрировал электрический потенциал. Классическое исследование параметров и механизма ПД проделано на гигантском аксоне кальмара с внутриклеточным раздражением и отведением внутриклеточного потенциала. В это нервное волокно (диаметром 0,5-1 мм) вводили на всю его длину два тончайших проволочных электрода. Один из них был раздражающим: через него в волокно подавали толчки тока того или иного направления, другой регистрировал электрический потенциал.

При подаче короткого и слабого толчка выходящего тока внутриклеточный электрод регистрировал кратковременное падение МП, по форме и силе соответствующее толчку тока, но со сглаженными передним и задним фронтами, что определяется емкостью мембраны. Это так называемый локальный потенциал. Локальным он называется потому, что и в экспериментальных, и в естественных уровнях он не распространяется далеко При подаче короткого и слабого толчка выходящего тока внутриклеточный электрод регистрировал кратковременное падение МП, по форме и силе соответствующее толчку тока, но со сглаженными передним и задним фронтами, что определяется емкостью мембраны. Это так называемый локальный потенциал. Локальным он называется потому, что и в экспериментальных, и в естественных уровнях он не распространяется далеко

. При усилении стимула и достижении порога раздражения, т.е. критического уровня деполяризации (КУД), возникает потенциал действия . При усилении стимула и достижении порога раздражения, т.е. критического уровня деполяризации (КУД), возникает потенциал действия

Перенос ионов через каналы биомембран
 и следовые потенциалы. В потенциале")
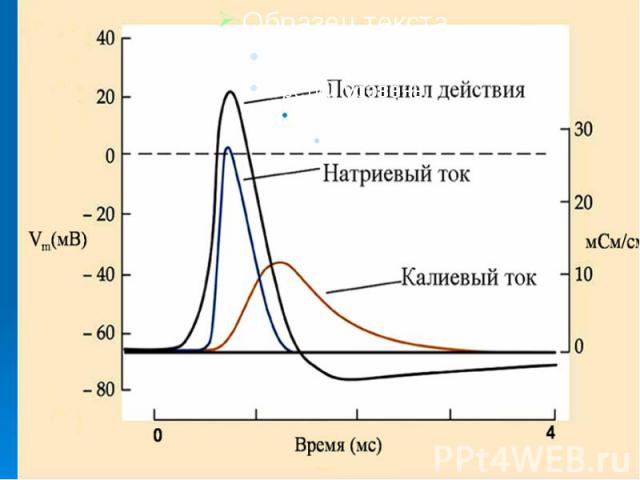
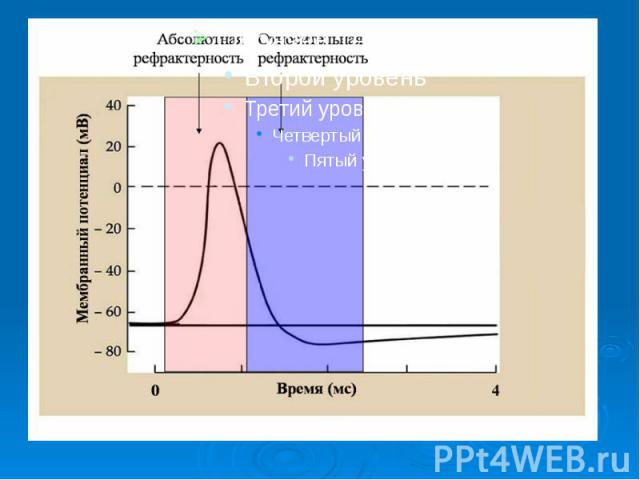
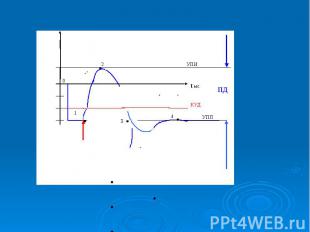
В потенциале действия различают пик (спайк) и следовые потенциалы. В потенциале действия различают пик (спайк) и следовые потенциалы. Пик ПД представляет собой кратковременную инверсию (изменение знака на положительный) внутриклеточного потенциала. Он имеет очень быструю восходящую фазу и несколько более медленный спад.

Общая длительность пика в данном объекте составляет около 3мс, амплитуда пика 120мВ, т.е. превышает МПП (90 мВ) на 30 мВ. Эту разницу называют овершутом или амплитудой потенциала инверсии. Общая длительность пика в данном объекте составляет около 3мс, амплитуда пика 120мВ, т.е. превышает МПП (90 мВ) на 30 мВ. Эту разницу называют овершутом или амплитудой потенциала инверсии.

Вслед за пиком ПД регистрируются значительно более слабые и длительные отрицательный и далее положительный следовой потенциалы Вслед за пиком ПД регистрируются значительно более слабые и длительные отрицательный и далее положительный следовой потенциалы

График изменения мембраного потенциала при разных воздействиях График изменения мембраного потенциала при разных воздействиях

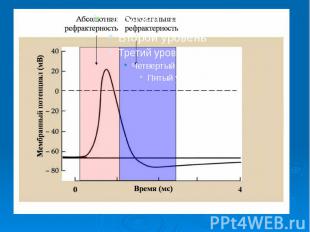
.График потенциала действия .График потенциала действия - возбуждающий импульс от генератора положительный (деполяризующий), но сдвиг мембранного потенциала аксона ниже некоторого порогового значения ( критического уровня деполяризации –КУД )-возникает локальный потенциал (1) . - амплитуда положительного.деполяризующего возбуждающего импульса приводит к сдвигу мембранного потенциала по значению выше порогового уровня, в мембране развивается генерация потенциала действия . 5. – при следовой гиперполяризации в течение 1-2 мс в мембране наблюдаются остаточные явления, во время которых мембрана остается рефракторной (невозбудимой).
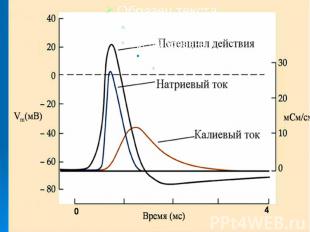

Потенциал действия имеет стандартные амплитуду и временные параметры, не зависящие от силы стимула, вызвавшего данный ПД (правило "все или ничего"). Потенциал действия имеет стандартные амплитуду и временные параметры, не зависящие от силы стимула, вызвавшего данный ПД (правило "все или ничего"). Потенциал действия - это электрический феномен, возникающий на плазматической мембране. Практически нормальный ПД возникает и в перфузируемом гигантском аксоне, лишенном аксоплазмы, при электрической стимуляции его мембраны.
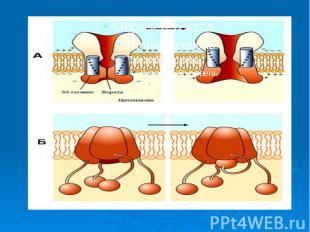

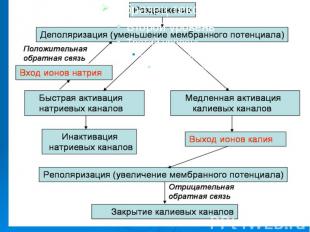
Причиной развития ПД является вызываемое критической деполяризацией мембраны открытие ее натриевых и калиевых каналов. Каналы, открываемые электрическим стимулом, называют потенциалзависимыми. Причиной развития ПД является вызываемое критической деполяризацией мембраны открытие ее натриевых и калиевых каналов. Каналы, открываемые электрическим стимулом, называют потенциалзависимыми.

Открытие потенциалзависимых каналов приводит к пассивному движению соответствующих ионов по их электрохимическим градиентам. Открытие потенциалзависимых каналов приводит к пассивному движению соответствующих ионов по их электрохимическим градиентам. Вход ионов в клетку обеспечивает восходящую фазу пика ПД, т.е. деполяризацию и инверсию потенциала на мембране, а несколько запаздывающий выход ионов участвует в создании нисходящей фазы пика - реполяризации.
. Связь разви")
При развитии пика ПД отношение становится (в покое оно составляет ). Связь развития пика ПД с током доказывается прямой зависимостью амплитуды ПД от электрохимического градиента на мембране и достоверным переходом меченого изотопа из среды в клетку при его возбуждении, причем в количестве, пропорциональном числу ПД. Связь нисходящей фазы ПД с током доказывается зависимостью хода этой фазы от электрохимического градиента на мембране. При развитии пика ПД отношение становится (в покое оно составляет ). Связь развития пика ПД с током доказывается прямой зависимостью амплитуды ПД от электрохимического градиента на мембране и достоверным переходом меченого изотопа из среды в клетку при его возбуждении, причем в количестве, пропорциональном числу ПД. Связь нисходящей фазы ПД с током доказывается зависимостью хода этой фазы от электрохимического градиента на мембране.
 в нервной клетке. Этот ответ облада")
Рассмотрим локальный подпороговый ответ (ЛО) в нервной клетке. Этот ответ обладает в основном тем же механизмом, что и ПД. Его восходящая фаза определяется входящим током Na, а нисходящая - выходящим током . Рассмотрим локальный подпороговый ответ (ЛО) в нервной клетке. Этот ответ обладает в основном тем же механизмом, что и ПД. Его восходящая фаза определяется входящим током Na, а нисходящая - выходящим током . Амплитуда ЛО пропорциональна силе раздражителя, а не стандартна, как у ПД, т.е. он не подчиняется правилу "все или ничего".

Рассчитаем равновесный электрохимический потенциал для натрия при возбуждении: Рассчитаем равновесный электрохимический потенциал для натрия при возбуждении: PNa+: PK+: PCl+=30:1:0,45 т.е. по сравнению с невозбужденным состоянием при возбуждении коэффициент проницаемости возрастает в 750 раз, т.к. , уравнение Гольдмана преобразуется в уравнение Нернста, и по нему можно рассчитать равновесный потенциал по натрию. Расчет по формуле Нернста.

В опытах было доказано, что этот равновесный потенциал по Na совпадает с амплитудным значением потенциала инверсии. ПИ всегда положителен, но меньше по абсолютной величине, чем потенциал В опытах было доказано, что этот равновесный потенциал по Na совпадает с амплитудным значением потенциала инверсии. ПИ всегда положителен, но меньше по абсолютной величине, чем потенциал покоя . ПД равен сумме ПП и ПИ

Характерные свойства потенциала действия: Наличие порогового значения деполяризующего потенциала. Закон "все или ничего", т.е., если деполяризующий потенциал больше порогового, развивается потенциал действия, амплитуда которого не зависит от амплитуды возбуждающего импульса и нет потенциала действия, если амплитуда деполяризующего потенциала меньше пороговой

Наличие периода рефрактерности, невозбудимости мембраны во время развития потенциала действия и остаточных явлений после снятия возбуждения. Наличие периода рефрактерности, невозбудимости мембраны во время развития потенциала действия и остаточных явлений после снятия возбуждения. Резкое уменьшение сопротивления мембраны в момент возбуждения (у аксона кальмара от 0,1 в покое до 0,0025 при возбуждении).

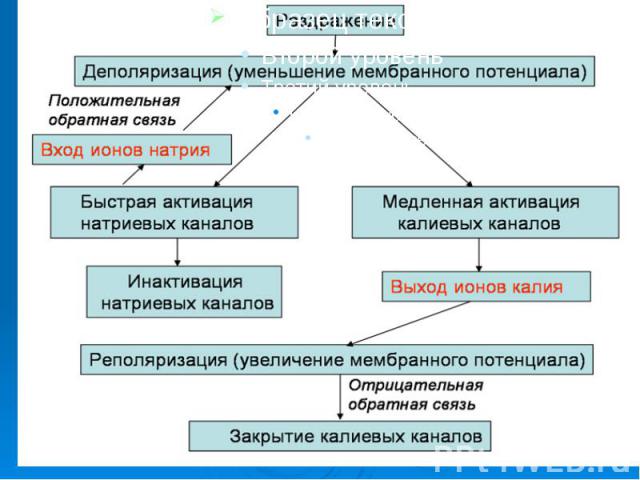
Итак, генерация потенциала действия в возбужденных мембранах возникает под влиянием различных факторов и сопровождается в первую очередь повышением проводимости клеточной мембраны для ионов натрия, входом их внутрь клетки, что приводит к деполяризации клеточной мембраны и появлению локального ответа. Итак, генерация потенциала действия в возбужденных мембранах возникает под влиянием различных факторов и сопровождается в первую очередь повышением проводимости клеточной мембраны для ионов натрия, входом их внутрь клетки, что приводит к деполяризации клеточной мембраны и появлению локального ответа.

. Этот процесс может достигнуть критического уровня деполяризации, после чего проводимость мембраны для натрия увеличивается до максимума, мембранный потенциал при этом приближается к натриевому равновесному потенциалу. Приблизительно через десять миллисекунд происходит инактивация натриевых каналов. . Этот процесс может достигнуть критического уровня деполяризации, после чего проводимость мембраны для натрия увеличивается до максимума, мембранный потенциал при этом приближается к натриевому равновесному потенциалу. Приблизительно через десять миллисекунд происходит инактивация натриевых каналов.

Увеличивается активация калиевых каналов, что приводит к увеличению выходящего калиевого тока, который в свою очередь вызывает реполяризацию и в дальнейшем восстановление потенциала покоя. Увеличивается активация калиевых каналов, что приводит к увеличению выходящего калиевого тока, который в свою очередь вызывает реполяризацию и в дальнейшем восстановление потенциала покоя.

Физиологическая роль ПД в потенциалзависимых клетках не ограничивается только возбудимостью клеточной мембраны и проведением нервного импульса. Возникновение ПД сопровождает целый комплекс информационных процессов, которые развиваются в ответ на стимуляцию на фоне ПД. Среди этих процессов следует назвать такие как: 1) изменение обмена веществ, 2) саморегуляция температуры и электрического импеданса, а так же других важных физиологических констант организма.

5. Распространение нервного импульса вдоль возбудимого волокна Если в каком-нибудь участке возбудимой мембраны сформировался потенциал действия, мембрана деполяризована, возбуждение распространяется на другие участки мембраны. Рассмотрим распространение возбуждения на примере передачи нервного импульса по аксону

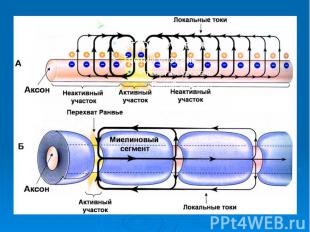
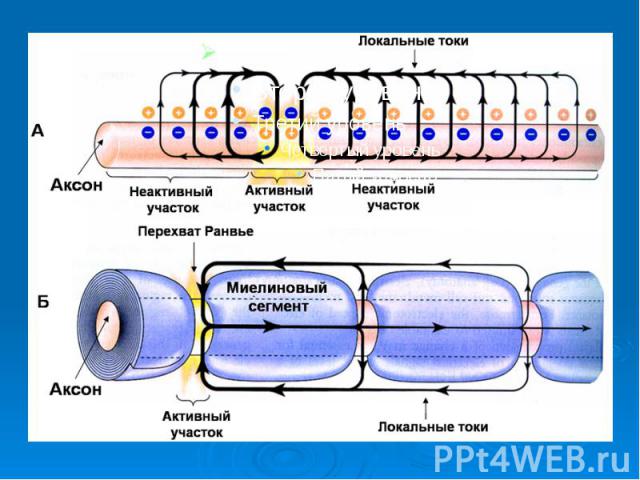
Локальные токи при распространении нервного импульса по нервному волокну

Положим, что один участок аксона находится в возбужденном состоянии, а соседний участок находится в покое. На границе таких зон возникают перепады мембранных потенциалов: плюс на возбужденном и минус на покоящемся участках. Надо учесть, что и аксоплазма и внеклеточная жидкость являются растворами электролитов, а следовательно, проводящими средами. Положим, что один участок аксона находится в возбужденном состоянии, а соседний участок находится в покое. На границе таких зон возникают перепады мембранных потенциалов: плюс на возбужденном и минус на покоящемся участках. Надо учесть, что и аксоплазма и внеклеточная жидкость являются растворами электролитов, а следовательно, проводящими средами.

Поэтому и в аксоплазме, и в окружающем растворе возникают локальные токи: между участками поверхности мембраны с большим потенциалом (положительно заряженными) и участками с меньшим потенциалом (отрицательно заряженными). Поэтому и в аксоплазме, и в окружающем растворе возникают локальные токи: между участками поверхности мембраны с большим потенциалом (положительно заряженными) и участками с меньшим потенциалом (отрицательно заряженными).

Локальные токи образуются и внутри аксона, и на наружной его поверхности. Локальные электрические токи приводят к повышению потенциала внутренней поверхности невозбужденного участка мембраны и к понижению наружного потенциала невозбужденного участка мембраны, оказавшегося по соседству с возбужденной зоной Локальные токи образуются и внутри аксона, и на наружной его поверхности. Локальные электрические токи приводят к повышению потенциала внутренней поверхности невозбужденного участка мембраны и к понижению наружного потенциала невозбужденного участка мембраны, оказавшегося по соседству с возбужденной зоной

Таким образом, в областях, близких к возбужденному участку, отрицательный потенциал покоя повышается и становится выше порогового значения. Под действием изменения мембранного потенциала открываются натриевые каналы и дальнейшее повышение происходит уже за счет потока ионов натрия через мембрану Таким образом, в областях, близких к возбужденному участку, отрицательный потенциал покоя повышается и становится выше порогового значения. Под действием изменения мембранного потенциала открываются натриевые каналы и дальнейшее повышение происходит уже за счет потока ионов натрия через мембрану

Происходит деполяризация мембраны, развивается потенциал действия. Затем возбуждение передается дальше на покоящиеся участки мембраны Происходит деполяризация мембраны, развивается потенциал действия. Затем возбуждение передается дальше на покоящиеся участки мембраны

Может возникнуть вопрос, почему возбуждение распространяется по аксону не в обе стороны от зоны, где возник нервный импульс, ведь локальные токи текут в обе стороны от возбужденного участка. Дело в том, что возбуждение может распространятся только в область мембраны, находящуюся в состоянии покоя, то есть в одну сторону от возбужденного участка аксона. В другую сторону нервный импульс не будет распространятся, так как области, через которые прошло возбуждение, некоторое время остаются невозбудимыми – рефракторными. Может возникнуть вопрос, почему возбуждение распространяется по аксону не в обе стороны от зоны, где возник нервный импульс, ведь локальные токи текут в обе стороны от возбужденного участка. Дело в том, что возбуждение может распространятся только в область мембраны, находящуюся в состоянии покоя, то есть в одну сторону от возбужденного участка аксона. В другую сторону нервный импульс не будет распространятся, так как области, через которые прошло возбуждение, некоторое время остаются невозбудимыми – рефракторными.

Эволюция животного мира привела к появлению нервных волокон, покрытых миелиновой оболочкой, которая представляет собой многомембранную систему. Миелин является изолятором, поэтому генерация ПД сосредоточена там, где миелиновая оболочка отсутствует, т.е. в перехватах Ранвье. Эволюция животного мира привела к появлению нервных волокон, покрытых миелиновой оболочкой, которая представляет собой многомембранную систему. Миелин является изолятором, поэтому генерация ПД сосредоточена там, где миелиновая оболочка отсутствует, т.е. в перехватах Ранвье. Аксоны позвоночных снабжены миелиновой оболочкой, которая увеличивает сопротивление мембраны и ее толщину

Возбуждение по миелинизированному волокну распространяется сальтаторно (скачкообразно) от одного перехвата Ранвье (участка, свободного от миелиновой оболочки) до другого. Возбуждение по миелинизированному волокну распространяется сальтаторно (скачкообразно) от одного перехвата Ранвье (участка, свободного от миелиновой оболочки) до другого.