
 Лекторы: А.П. Перевозчиков, М.А. Кулакова, Д. А. Могиленко, …")




. Они отражают главные планы строения всех многоклеточных животных организмов, образовавшихся в раннем кембрии. Вероятно, это максимальное число…")




, созданная к середине 20-го века (Добжанский, Симпсон, Майр и др.) базировалась в основном на достижениях популяционной генетики, учитывающей мутации единичных генов, тестируемые у взрослых особей. Она могла об…")

 и его значение в создании EVO/DEVO Тройной подход к созданию эволюционной биологии (палеонтологические, анатомические и эмбриологические доказательства) Теория происхождения многоклеточных (гастрея) Биогенетический закон Спра…")



, но не обязательно гомологичных структур")
 гены по ходу эволюции могут расщирять (менять) функцию за счёт явления коопции, и приобретать тем самым новые функции")
 биохимических реакций, включая каскады путей сигналинга. Регуляторные гены также как и структурные могут быть…")




 и у человека (NF-kappaB).")

. Желтым и коричневым цветом попарно отмечены ортологичные факторы. Пронейральные гены: Achaete-Scute/ Mash1,2. Гомеотические гены -Hom/Hox, гомеобокс-содержащие ген…")
, направления осей роста конечности (передне-задняя, дорсо-вентральная, проксимо-дистальная)")
 курицы и дрозофилы")




 и морфологических (справа) критериев")





 и наименее специфичные в отношении лигандов")





 белка, которого ген кодирует Функция белка –понятие отражающее эволюционный аспект его свойств и, следовательно, имеет меняющееся со временем значение. Нельзя также не учитывать и субъективизм по…")
. В отличие …")
Презентация на тему: Эволюция животных

Лекция 1 Эволюционная биология развития Роль процессов развития в эволюции. Новый синтез идей в понимании процессов микро- и макроэволюции -EvoDevo. (автор курса –проф. А. П. Перевозчиков) Лекторы: А.П. Перевозчиков, М.А. Кулакова, Д. А. Могиленко, Е. Б. Малашичев, Р. П. Костюченко В этом курсе (там где особо не указано) используются материалы из учебного пособия S. Gilbert “Developmental Biology” 2003, “Sinauer” 900igr.net

From W. Gehring’ Lecture

Эволюция и рост биоразнообразия – связанные понятия

Царства живых организмов Protista unicellular eukaryotes and algae Plantae -multicellular photosynthetic cell walls Fungi -multicellular filamentous heterotrophic cell walls Monera -prokaryotes Animalia 250,000 sp. 60,000 sp. 5,000 sp. 5,000 sp. 1,000,000+ sp. ? 5 Kingdom Classification

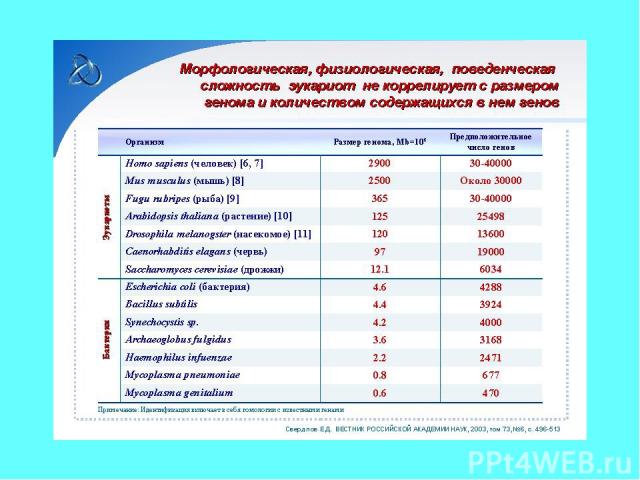
Многие типы современных многоклеточных животных зародились в раннем кембрии

Существует примерно 35 архетипов организации животных (в, основном, соответствующих числу типов животных). Они отражают главные планы строения всех многоклеточных животных организмов, образовавшихся в раннем кембрии. Вероятно, это максимальное число всех способных к возникновению путём эволюции вариантов (bauplan) организмов.

Эволюционная теория должна помочь объяснению явления биоразнообразия

В девятнадцатом веке существовали два диаметрально противоположных взгляда на происхождение видов. Один из них, наиболее ярко выраженный Жоржем Кювье и Чарльзом Беллом, фокусировался на различиях между видами, позволяющими каждому из них адаптироваться к тому окружению, в котором он живет. В свете этих представлений рука человека, плавник тюленя, крылья птицы и летучей мыши представали замечательными изобретениями Творца, дающими этим животным возможность приспособиться к "условиям существования." В соответствии с другим взглядом, наиболее энергичными сторонниками которого были Этьен Жоффруа Сент-Илер и Ричард Оуэн, ключевое значение принадлежало “единству типа” (чертам сходства между организмами, которые Оуэн называл “гомологиями”). Рука человека, плавник тюленя, крылья птицы и летучей мыши представлялись ими как модификации одного и того же основного плана. Поиски этого плана могли привести к форме, по которой Творец создал этих животных. Адаптациям они отводили второстепенную роль.

«Позвоночное – это омар, перевернутый на спину» - Этьен Жоффруа Сент-Илер

Дарвин отдал должное этим дебатам, когда в 1859 году написал: ”Общепринято, что строение всех органических существ подчинено двум великим законам – Единства Типа и Единства Условий Существования”. Далее Дарвин пишет, что его теория могла бы объяснить единство типа происхождением от общего предка. Изменения, создающие замечательные адаптации к условиям существования, может объяснить теория естественного отбора. Свою концепцию Дарвин назвал ”происхождением с модификацией”.
, созданная к середине 20-го века (Добжанс")
Теория эволюции («современный синтез»), созданная к середине 20-го века (Добжанский, Симпсон, Майр и др.) базировалась в основном на достижениях популяционной генетики, учитывающей мутации единичных генов, тестируемые у взрослых особей. Она могла объяснить лишь процессы микроэволюции, ведущие к видообразованию, но не смогла найти механизмов макроэволюции, ведущих к крупным перестройкам и к появлению крупных таксонов - классов и типов. Эмбриологические результаты были вычеркнуты из цепочки доказательств

 и его значение в создании EVO/DEVO Тройной подход к созд")
Э. Геккель ( 1843-1919) и его значение в создании EVO/DEVO Тройной подход к созданию эволюционной биологии (палеонтологические, анатомические и эмбриологические доказательства) Теория происхождения многоклеточных (гастрея) Биогенетический закон Справедливая критика закона, вместе с тем, ведет к отрицанию единообразия (морфологических) форм и путей развития Что же лежит в основе эволюционной идеи (что её консолидирует)?

Вопросы макроэволюции с точки зрения теории «нового синтеза» не могли быть решены Вероятно, к крупным изменениям в структуре и физиологии, совместимым с жизнью организм может приспособиться лишь в процессе раннего развития. Генетические основы: значительные перестройки могут вызываться хромосомными перестройками, мутациями генов-селекторов, высокой активностью траснпозонов и мутациями генов системного уровня организации (гормональной, иммунной, нервной и др. систем)

Вероятно, к крупным изменениям в структуре и физиологии, совместимым с жизнью, организм может приспособиться лишь в процессе раннего развития. Значительные перестройки вызываются мутациями генов-селекторов, высокой активностью транспозонов и мутациями генов системного уровня организации (гормональной, иммунной, нервной и т.п.)

НАЗРЕЛА НЕОБХОДИМОСТЬ НОВОГО «НОВОГО СИНТЕЗА» ИДЕЙ ЭВОЛЮЦИИ, то есть теперь уже новых подходов к эволюции с использованием данных геномики и протеомики, эмбриогенетики, молекулярной биологии развития и палеонтологии, биоинформатики и других подходов. Часть идей нового синтеза отражена в формирующейся новой науке : Evolution and Development (Evo/Devo). Она может объяснить быстрое возникновение макро-инноваций
, н")
Гомологичные гены отвечают за образование функционально сходных (аналогичных), но не обязательно гомологичных структур
 гены по ходу эволюции могут расщирять (менять) функц")
Гомологичные (ортологичные) гены по ходу эволюции могут расщирять (менять) функцию за счёт явления коопции, и приобретать тем самым новые функции

Решение парадокса эволюции форм лежит на уровне молекулярной биологии развития Кроме ортологичных генов, сохраняются целые цепочки (звенья) биохимических реакций, включая каскады путей сигналинга. Регуляторные гены также как и структурные могут быть ортологичны Каскады регуляций генетических программ, каскады сигнальных цепочек проявляют удивительный консерватизм и являются основой эволюционных преобразований

Число генов в геномах многоклеточных организмов хотя и увеличивается в эволюционом ряду от низших к высшим, но всё же не настолько, чтобы объяснить все эволюционные усложнения организации. Вероятнее, что многие гены высших животных гомологичны генам низших и могут контролировать сходные функции, приобретая одновременно и новые. Такие гены называются ортологичными. Гомологичные гены в геноме животных одного и того же вида называются паралогичными. Главный путь усложнения геномов - усложнение регуляторных механизмов экспрессии генов

Консерватизм сигнальной цепочки, передающей сигнал через тирозинкиназый рецептор

Wnt-сигнальный каскад у различных организмов

Консерватизм сигнальной цепочки, действующей через Toll- рецепторы у Drosophila и млекопитающих. Сигнальная цепочка, действующая через Toll-рецептор, используется для запуска воспалительного ответа у млекопитающих и для активации антифунгального действия у взрослых мух. Гомологичные белки отмечены одинаковым цветом

Использование гомологичных компонентов сигнальной цепочки, включая ТФ: гомологичные Dorsal/Dif /Relish, в иммунных реакциях у дрозофилы (антифунгальный и антибактериальный ответы) и у человека (NF-kappaB).

Консерватизм BMP/Chordin -сигналинга и использование его в различных формообразовательных процессах

Гомология путей спецификации нейральной эктодермы у дрозофилы и позвоночных(Xenopus). Желтым и коричневым цветом попарно отмечены ортологичные факторы. Пронейральные гены: Achaete-Scute/ Mash1,2. Гомеотические гены -Hom/Hox, гомеобокс-содержащие гены Lim1,3, Isl/Lim1,3, Isl1,2

Кости конечности позвоночного. Расположение костей в скелете куриного крыла (стилоподий, зигоподий, аутоподий), направления осей роста конечности (передне-задняя, дорсо-вентральная, проксимо-дистальная)

Гомология процессов образования передне-задней оси негомологичных конечностей (крыльев) курицы и дрозофилы

Высокая гомология регуляторов, контролирующих формирование крыла курицы и дрозофилы

Роль гомеозисных генов в спецификации нервной системы вдоль передне-задней оси. Градиент ретиноевой кислоты в передне-заднем направлении модифицирует активность Hox-генов.

Классическое представление филогении животных Diploblast Triploblast and Bilaterian Coelomate animals Acoelomate and Pseudo-coelomate animals No organised tissue layers

Филогения на основании анализа генетической последовательности 18S рибосомной РНК

Эволюционные древа животных, построенные на основании молекулярно-генетических (слева) и морфологических (справа) критериев

Генетическая перекрываемость между различными царствами животных

Предосторожности от расширенного толкования термина «гомологичные гены» Существованием гомологичных генов не всегда можно объяснить преемственность функций, которые контролируются этими генами у различных животных

Биосинтез кортикальных и половых стероидов

Эволюционное родство хордовых животных

Стероидные гормоны, родственные кортикальным и половым гормонам позвоночных найдены у некоторых беспозвоночных, но не являются у них облигатными формами: они могут обнаружены у некоторых ракообразных и моллюсков. Возможно, синтезируются у ланцетника! Гомологи их ядерных рецепторов обнаружены у моллюска Aplysia californica, отсутствуют у асцидии и морского ежа и обнаружены у миксины и миноги. Есть ли они у ланцетника – вопрос!

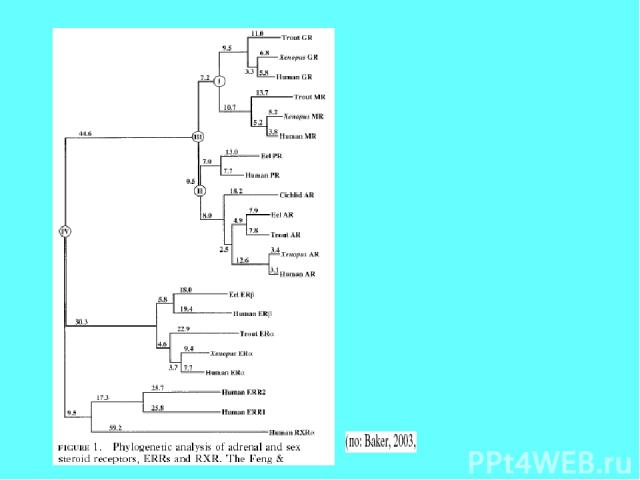
Эволюционное исследование стероидных рецепторов хордовых показало, что эстрогеновые рецепторы наиболее древние по сравнению со всеми другими (ближе всего к ретиноидным рецепторам) и наименее специфичные в отношении лигандов



Две модели действия эстрогена: классическая и «быстрого ответа»


Предосторожности от расширенного толкования термина «гомологичные гены» Существованием гомологичных генов не всегда можно объяснить преемственность функций, которые контролируются этими генами у различных животных
 белка, которого ген кодиру")
Фунция гена означает биологическое свойство (функцию) белка, которого ген кодирует Функция белка –понятие отражающее эволюционный аспект его свойств и, следовательно, имеет меняющееся со временем значение. Нельзя также не учитывать и субъективизм понятия функции гена, зависящий от исследователя Далее примеры:

У гена эстрогенового рецептора млекопитающих есть родственники, которые можно рассматривать как паралоги, хотя они имеют всего лишь 35% идентичности и 60% положительных совпадений по нуклеотидным последовательностям (цит. по Baker, 2008). В отличие от генов эстрогенового рецептора ER (estrogen receptor)- ERa и ERb, их гены называются ERR (estrogen-receptor-related)- ERRa и ERRg соответственно; кодируемые ими белки не способны связываться со стероидами и по сути являются орфановыми ядерными рецепторами. Функции их не известны, по-видимому, они играют роль в остеогенезе. Благодаря секвенированию геномов многих позвоночных и беспозвоночных животных удалось проследить родственные связи между генами ER и ERR позвоночных и беспозвоночных животных. Гены, аналогичные генам ERR позвоночных идентифицируются в геномах , практически, всех крупных таксонов беспозвоночных и найдены даже у весьма примитивного животного организма Trichoplax Можно утверждать, что эволюционно гены ER/ERR имели общего предкового предшественника, который появился у эволюционно древних беспозвоночных (по меньшей мере, у Bilateria). Кодируемые этими генами белки не выполняли функции связывания стероидов, а выполняли какие-то другие функции.











