
 мышечные ткани: 2.1. Скелетная мышечная ткань. 2.2. Сердечная мышечная ткань. 3. Гладкая мышечная ткань. 4. Гистофизиология сокращения мышц. 5. Регенера…")

 мышечные ткани делятся на 5 типов: мезенхимные (мышечные ткани внутренних органов, кроме сердца); эпидермальные (миоэпителиоциты желез – производные эктодермы); нейр…")


, а в центральной его части локализ…")
 и миозиновыми (толстыми) миофиламентами, котор…")
, представляющая собой сетевидную структуру цитоскелета, образованную фибриллярным белком α-актинином. К узлам этой сети актиновые филаменты фиксируются с помощью белков небулина, виментина и …")
, переходные, проводящие и секреторные. Все типы кардиомиоцитов покрыты базальной…")
 кардиомиоциты имеют форму цилиндра длиной около 100-150 мкм и диаметром до 20 мкм. Они содержат одно, реже два ядра, которые располагаются по центру клетки, а вокруг ядер группами локализуются миофибриллы (поля Конгейма).")


 кардиомиоциты – задаватели ритма. Они воспринимают управляющие сигналы от нервных волокон и в соответствии с этим изменяют ритм сократительной деятельности. Пейсмекеры передают управляющие сигналы на п…")












 ↑ (с…")
. Погибающие кардиомиоциты не восстанавл…")
, и…")

Презентация на тему: Мышечные ткани

Лекция Мышечные ткани План 1. Классификация мышечных тканей. 2. Поперечнополосатые (исчерченные) мышечные ткани: 2.1. Скелетная мышечная ткань. 2.2. Сердечная мышечная ткань. 3. Гладкая мышечная ткань. 4. Гистофизиология сокращения мышц. 5. Регенерация мышечных тканей. 6. Строение мышцы как органа. Составитель – профессор Н.П. Барсуков Симферополь 2008

В группу мышечных тканей входят различные как по строению, так и по происхождению ткани, общим для которых является способность активно сокращаться. Они обеспечивают перемещение тела в пространстве и его частей относительно друг друга.

Классификация мышечных тканей 1. По происхождению (гистогенетическая классификация) мышечные ткани делятся на 5 типов: мезенхимные (мышечные ткани внутренних органов, кроме сердца); эпидермальные (миоэпителиоциты желез – производные эктодермы); нейральные (из нервной трубки); целомические (из миоэпикардиальной пластинки висцерального листка несегментированной мезодермы); соматические (из миотомов). 2. По морфофункциональным особенностям мышечные ткани делятся на поперечнополосатые (исчерченные) и гладкие.

Морфология мышечных тканей Характерной морфологической особенностью всех типов мышечных тканей является удлинённая форма их структурных компонентов, которые содержат специальные органеллы – миофибриллы или миофиламенты, состоящие из сократительных белков.

Скелетная мышечная ткань Структурной единицей скелетной мышечной ткани является мышечное волокно, которое состоит из миосимпласта и миосателлитоцитов, покрытых общей базальной мембраной. Базальная мембрана вместе с плазмолеммой образуют сарколемму.

Длина миосимпласта колеблется от нескольких микрометров до нескольких сантиметров, а диаметр составляет 50-100 мкм. По периферии миосимпласта располагаются ядра (от нескольких десятков до нескольких десятков тысяч), а в центральной его части локализуются миофибриллы.

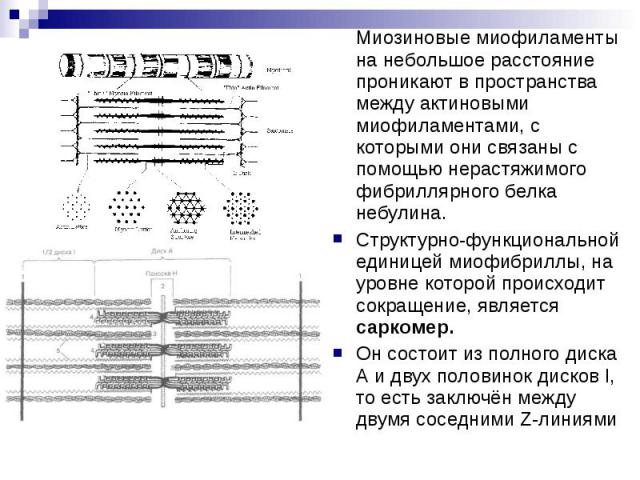
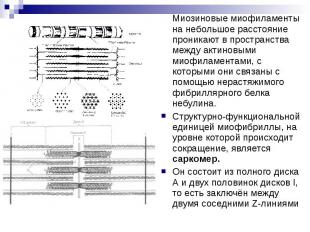
Скелетная мышечная ткань. ЭГ. Миофибриллы строго ориентированы вдоль волокна и состоят из чередующихся светлых и темных полос, или I-дисков и А-дисков, образованных, соответственно, актиновыми (тонкими) и миозиновыми (толстыми) миофиламентами, которые располагаются параллельно друг другу. Благодаря строгой ориентации миофибрилл мышечные волокна имеют поперечную исчерченность.
, представляющая собой")
В середине каждого I-диска находится телофрагма (Z-линия), представляющая собой сетевидную структуру цитоскелета, образованную фибриллярным белком α-актинином. К узлам этой сети актиновые филаменты фиксируются с помощью белков небулина, виментина и десмина. Миозиновые филаменты, образующие А-диски, в своей средней части также скрепляются с цитоскелетом - мезофрагмой, представляющей сетевидную структуру, состоящую из белка миомезина. Это место называется М-линией. Миозиновые филаменты не доходят своими концами до Z-линий, но связаны с ними с помощью растяжимого белка титина.

Сердечная мышечная ткань Её труктурной единицей являются клетки – кардиомиоциты. Различают 5 их типов: сократительные, или типичные и атипичные: синусные (пейсмекерные), переходные, проводящие и секреторные. Все типы кардиомиоцитов покрыты базальной мембраной.
 кардиомиоциты имеют форму цилиндра длиной около 100-15")
Типичные (сократительные) кардиомиоциты имеют форму цилиндра длиной около 100-150 мкм и диаметром до 20 мкм. Они содержат одно, реже два ядра, которые располагаются по центру клетки, а вокруг ядер группами локализуются миофибриллы (поля Конгейма).

Сократительные кардиомиоциты соединяются торец в торец, образуя функциональные мышечные волокна. В области соединений кардиомиоцитов чётко выявляются вставочные диски. Строение миофибрилл такое же, как и в скелетной мышечной ткани.

Схема ультрамикроскопического строения сократительных кардиомиоцитов
 кардиомиоциты – задаватели ритма")
Атипичные кардиомиоциты Синусные (пейсмекерные) кардиомиоциты – задаватели ритма. Они воспринимают управляющие сигналы от нервных волокон и в соответствии с этим изменяют ритм сократительной деятельности. Пейсмекеры передают управляющие сигналы на переходные кардиомиоциты, от которых сигнал поступает на проводящие (пучок Гисса, волокна Пуркинье) и далее – на сократительные кардиомиоциты.

Секреторные кардиомиоциты относятся к гормонопродуцирующим клеткам. Они вырабатывают натрийуретический фактор, который участвует в регуляции мочеобразования, а также кровяного давления и др. процессов.

Морфологические особенности атипичных кардиомиоцитов Они имеют овальную форму, крупнее диаметром в 2-3 раза, чем типичные, миофибрилл у них соответственно меньше, которые располагаются неупорядоченно, поэтому поперечная исчерченность не выражена В них больший объём цитоплазмы, которая при окрашивании гораздо светлее, ядро располагается эксцентрично. В них слабо представлены органеллы общего значения: ЭПС, митохондрии, отсутствуют Т-трубочки. При гистохимических исследованиях в них выявляется больше включений гликогена, высокая активность ЛДГ в то время как в типичных – меньше гликогена и высокая активность СДГ.

Гладкая мышечная ткань мезенхимного типа Её структурной единицей является миоцит, имеющий веретеновидную форму, ядро в нём удлинённое, локализуется по центру клетки. Длина миоцитов колеблется в пределах 20-500 мкм, а диаметр в области брюшка – всего лишь 5-8 мкм.

Ультраструктура миоцита

Гладкая мышечная ткань эпидермального происхождения

Гладкая мышечная ткань нейрального происхождения Развивается из нейрального зачатка внутреннего слоя глазного бокала. Миоциты локализуются в эпителии радужки, имеют отростки, в которых находится сократительные миофиламенты. Миоциты образуют 2 мышцы – суживающую и расширяющую зрачок.

Гистофизиология сокращения исчерченных мышц. Cкелетные мышцы богато иннервированы

Сигнал о начале сокращения исчерченной мышечной ткани задаётся центральной нервной системой. Он вызывает волну деполяризации плазмолеммы, которая в виде потенциала действия передаётся на мембрану Т-трубочек и далее на мембрану АЭC, вследствие чего из её цистерн высвобождаются ионы кальция, инициирующие взаимодействие актиновых и миозиновых миофиламентов, то есть сокращение. После прекращения сигнала ионы кальция снова аккумулируются в АЭС и сокращение прекращается.

В процессе сокращения поперечнополосатых мышц длина актиновых и миозиновых филаментов не изменяется, а происходит их смещение относительно друг друга: миозиновые нити вдвигаются в пространства между актиновыми, а актиновые - между миозиновыми; в результате этого: ширина I-диска и H-полоски А-диска уменьшается, в то же время ширина диска А не изменяется, но длина саркомера укорачивается. 1 – актин; 2 – головка миозина.

Сократительный аппарат гладкомышечных клеток представлен филаментами актина, образующими трёхмерную сеть, рядом с которой располагаются мономеры миозина.

В отличие от поперечнополосатых мышц, в которых миофибриллы существуют постоянно, в гладких мышцах они образуются только в момент сокращения, которое происходит вследствие поступления сигнала от нервных клеток.

Под воздействием медиатора в плазмолемме миоцитов образуются кавеолы, в которые путём эндоцитоза поступают ионы кальция, вызывающие полимеризацию мономеров миозина и его взаимодействие с актиновыми филаментами. 2 – плотные тельца; 8 – актиновые и 9 – миозиновые миофиламенты.

Актиновые филаменты одним своим концом с помощью сшивающих белков прикрепляются к специальным участкам внутренней поверхности плазмолеммы, а другим - к миозину. Миозиновые филаменты прикрепляются к специальным местам в цитозоле клетки (нексусы) ↑ (см. предыдущий слайд).

Регенерация мышечных тканей Поперечнополосатая скелктная мышечная ткань регенерирует за счёт миосателлитоцитов. В сердечной исчерченной мышечной ткани возможна внутриклеточная регенерация (рабочая гипертрофия). Погибающие кардиомиоциты не восстанавливаются. Гладкая мышечная ткань также регенерирует на клеточном уровне (рабочая гипертрофия).

Строение мышцы как органа Мышечные ткани образуют органы или входят в состав других органов. В том и ином случае они тесно взаимодействуют с волокнистой соединительной тканью, которая в виде прослоек окружает мышечные волокна и клетки (эндомизий), их группы (перимизий) и мышцу как орган (эпимизий). В соединительной ткани проходят сосуды и нервы.

БЛАГОДАРЮ ЗА ВНИМАНИЕ

»")
»")


»")





»")