









 с соответствующими моторными молекулами. Р - микротрубочки реснички.Молекулы динеина прикреплены хвостами (линии) и головками (черные кружки) к соседним микротрубочкам. Перемещение головок вызывает сгибание об…")
, центросфера, цетроплазма или клеточный центр — главный центр организации микротрубочек (ЦОМТ) и регулятор хода клеточного цикла в клетках эукариот. Впервые обнаружена в 1888 году Теодором Бовери, который назвал её «ос…")

, а Э.ван Бенеден - в интерфазной клетке.")
![Наряду с центросомами, также в конце XIX в., были описаны органеллы, лежащие у основания специализированных клеточных образований - ресничек и жгутиков; эти органеллы получили название кинетосом, или базальных телец [2, 3]. Авторы, Л.Хеннеги и М.Лег…](/images/1469/45072/640/img13.jpg "Наряду с центросомами, также в конце XIX в., были описаны органеллы, лежащие у основания специализированных клеточных образований - ресничек и жгутиков; эти органеллы получили название кинетосом, или базальных телец [2, 3]. Авторы, Л.Хеннеги и М.Лег…")

![Рис. 3. Ультраструктура центросомы в интерфазной клетке млекопитающих на последовательных серийных срезах [19].](/images/1469/45072/640/img15.jpg "Рис. 3. Ультраструктура центросомы в интерфазной клетке млекопитающих на последовательных серийных срезах [19].")


, центросому (зеленый цвет) и ДНК (синий цвет). Положение центросом…")
![Микротрубочки и связь с клиникойСиндром неподвижных ресничек, или синдром первичной дискинезии ресничек (syndrome of immobile cilia or immotile cilia syndrome) - гетерогенная группа заболеваний (МIМ:242630, 242670, 242680, 244400) [14, 38]. Синдром …](/images/1469/45072/640/img19.jpg "Микротрубочки и связь с клиникойСиндром неподвижных ресничек, или синдром первичной дискинезии ресничек (syndrome of immobile cilia or immotile cilia syndrome) - гетерогенная группа заболеваний (МIМ:242630, 242670, 242680, 244400) [14, 38]. Синдром …")

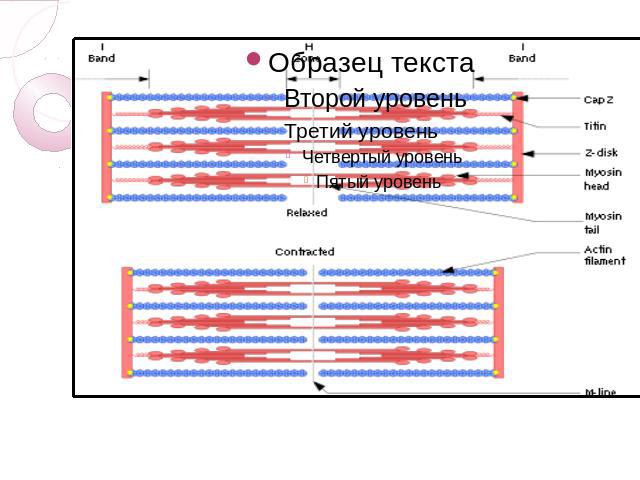
. Миофибриллы — структуры толщиной 1 — 2 мкм, состоящие из саркомеров — структ…")

. Скорость скольжения миозина по актину ≈15 мкм/сек. В миозиновом филаменте много (около 500) молекул миозина и, следовательно, при со…")

.")


Презентация на тему: Двигательные органеллы клетки

Карагандинский государственный медицинский университетКафедра молекулярной биологии и медицинской генетики СРС на тему: «Двигательные органеллы клетки»

МикротрубочкиМикротрубочки представляют собой полые внутри цилиндры диаметром 25 нм. Длина их может быть от нескольких микрометров до, вероятно, нескольких миллиметров в аксонах нервных клеток. Их стенка образована димерами тубулина. Микротрубочки, подобно актиновым микрофиламентам, полярны: на одном конце происходит самосборка микротрубочки, на другом — разборка. В клетках микротрубочки играют роль структурных компонентов и участвуют во многих клеточных процессах, включая митоз, цитокинез и везикулярный транспорт.

В образовании микротрубочки выделяют три фазы: Замедленная фаза, или нуклеация. Это этап зарождения микротрубочки, когда молекулы тубулина начинают соединяться в более крупные образования. Такое соединение происходит медленнее, чем присоединение тубулина к уже собранной микротрубочке, поэтому фаза и называется замедленной;Фаза полимеризации, или элонгация. Если концентрация свободного тубулина высока, его полимеризация происходит быстрее, чем деполимеризация на минус-конце, за счет чего микротрубочка удлиняется. По мере её роста концентрация тубулина падает до критической и скорость роста замедляется вплоть до вступления в следующую фазу;Фаза стабильного состояния. Деполимеризация уравновешивает полимеризацию, и рост микротрубочки останавливается.

Динамическая нестабильность

ФункцияМикротрубочки в клетке используются в качестве «рельсов» для транспортировки частиц. По их поверхности могут перемещаться мембранные пузырьки и митохондрии. Транспортировку по микротрубочкам осуществляют белки, называемые моторными. Это высокомолекулярные соединения, состоящие из двух тяжёлых (массой около 300 кДа) и нескольких лёгких цепей. В тяжёлых цепях выделяют головной и хвостовой домены. Два головных домена связываются с микротрубочками и являются собственно двигателями, а хвостовые — связываются с органеллами и другими внутриклеточными образованиями, подлежащими транспортировке.

СтроениеМикротрубочки — это структуры, в которых 13 тубулиновых α-/β-гетеродимеров уложены по окружности полого цилиндра. Внешний диаметр цилиндра около 25 нм, внутренний — около 15.Один из концов микротрубочки, называемый плюс-концом, постоянно присоединяет к себе свободный тубулин. От противоположного конца — минус-конца — тубулиновые единицы отщепляются.

Строение микротрубочки

ЦентросомаЦОМТ - центросомы, из которых растет митотическое веретено и "звезды" микротрубочек во многих клетках, а также базальные тельца, из которых растут микротрубочки жгутиков и ресничек. Замечательное свойство этих центров, что они способны репродуцироваться: новый центр вырастает рядом со старым и затем "материнский" и дочерний центры расходятся. Долго искали в центрах ДНК, но не нашли. Удвоение центров, видимо, имеет совсем особый механизм, отличный от удвоения ДНК, но природу его мы еще не знаем.Как уже говорилось, микротрубочки разных структур сильно различаются по стабильности

Структуры, образуемые системой микротрубочек и организующих их центров.А - параллельные микротрубочки ресничек, растущие от базальных телец к поверхности клетки.Б - микротрубочки митотического веретена, растущие навстречу друг другу от двух центров, расположенных на полюсах веретена. Некоторые из микротрубочек прикрепляются плюс-концами к особым участкам (кинетохорам) хромосом. На схеме две хромосомы в метафазе, то есть до начала расхождения к полюсам
 с соответствующими моторными молекулами.")
Взаимодействия микротрубочек (цилиндры) с соответствующими моторными молекулами. Р - микротрубочки реснички.Молекулы динеина прикреплены хвостами (линии) и головками (черные кружки) к соседним микротрубочкам. Перемещение головок вызывает сгибание обеих микротрубочек. К, Д - движения органелл (большие круги) при помощи молекул кинезина (К) или динеина (Д) вдоль микротрубочки в противоположных направлениях.
, центросфера, цетроплазма или клеточный центр — г")
Центросома (греч. soma — тело), центросфера, цетроплазма или клеточный центр — главный центр организации микротрубочек (ЦОМТ) и регулятор хода клеточного цикла в клетках эукариот. Впервые обнаружена в 1888 году Теодором Бовери, который назвал её «особым органом клеточного деления». Хотя центросома играет важнейшую роль в клеточном делении, недавно было показано, что она не является необходимой. В подавляющем большинстве случаев в клетке в норме присутствует только одна центросома. Аномальное увеличение числа центросом характерно для раковых клеток. Более одной центросомы в норме характерно для некоторых полиэнергидных простейших и для синцитиальных структур.

Центросома

Такой увидели центросому в 1887 г. ее первооткрыватели: Т.Бовери описал ее в полюсах митотического веретена (слева), а Э.ван Бенеден - в интерфазной клетке.

Наряду с центросомами, также в конце XIX в., были описаны органеллы, лежащие у основания специализированных клеточных образований - ресничек и жгутиков; эти органеллы получили название кинетосом, или базальных телец [2, 3]. Авторы, Л.Хеннеги и М.Легоссек, наблюдали взаимный переход базальных телец и центросом и в 1898 г. выдвинули гипотезу о гомологии этих клеточных органелл, которая впоследствии получила экспериментальное подтверждение (рис.2).

Рис.2. Формирование полюсов веретена деления из базальных телец в сперматоцитах Bombyx mori [ах Bombyx mori [

Рис. 3. Ультраструктура центросомы в интерфазной клетке млекопитающих на последовательных серийных срезах [19].

Упрощенная схема строения центросомы в интерфазных клетках млекопитающих в середине S-фазы клеточного цикла.

Центросома, окруженная комплексом Гольджи. На ультратонком срезе располагается одна центриоль из пары

Центросома и система микротрубочек в профазной, метафазной и интерфазной клетках. Световая микроскопия. Тройное иммунофлуоресцентное окрашивание выявляет микротрубочки (красный цвет), центросому (зеленый цвет) и ДНК (синий цвет). Положение центросом показано стрелками.

Микротрубочки и связь с клиникойСиндром неподвижных ресничек, или синдром первичной дискинезии ресничек (syndrome of immobile cilia or immotile cilia syndrome) - гетерогенная группа заболеваний (МIМ:242630, 242670, 242680, 244400) [14, 38]. Синдром Картагенера рассматривают как классический вариант первичной дискинезии ресничек [37], характерным признаком которого, помимо бронхолегочных заболеваний и поражения носоглотки, является обратное расположение внутренних органов (situs inversus). Впервые эта патология описана А.К. Зивертом в 1902 г. в журнале «Русский врач» [46].Клиника: синдром характеризуется повторяющимися инфекциями дыхательных путей, хроническим бронхитом, пансинуситом, отитом, бесплодием (мужским и женским). У пациентов развиваются аносмия, полипы носовой полости, умеренное снижение слуха по проводящему типу [4, 5, 10].Развитие бронхолегочных инфекций и, нередко, бесплодия связано с нарушением двигательной активности ресничек мерцательного эпителия слизистой оболочки дыхательных путей и изменением числа синглетов и дуплетов микротрубочек в аксонеме фимбрий воронки и полости маточных труб, в аксонеме ресничек клеток реснитчатого эпителия дыхательных путей и жгутиков сперматозоидов.

Мышечное сокращение — реакция мышечных клеток на воздействие нейромедиатора, реже гормона, проявляющаяся в уменьшении длины клетки. Эта жизненно важная функция организма, связанная с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами

Основой всех типов мышечного сокращения служит взаимодействие актина и миозина. В скелетных мышцах за сокращение отвечают миофибриллы (примерно две трети сухого веса мышц). Миофибриллы — структуры толщиной 1 — 2 мкм, состоящие из саркомеров — структур длиной около 2,5 мкм, состоящих из актиновых и миозиновых (тонких и толстых) филаментов и Z-дисков, соединённых с актиновыми филаментами. Сокращение происходит при увеличении концентрации в цитоплазме ионов Ca2+ в результате скольжения миозиновых филаментов относительно актиновых. Источником энергии сокращения служит АТФ. КПД мышечной клетки около 50 %.

Скольжение миозина относительно актинаГоловки миозина расщепляют АТФ и за счет высвобождающейся энергии меняют конформацию, скользя по актиновым филаментам. Цикл можно разделить на 4 стадии:Свободная головка миозина связывается с АТФ и гидролизует его до АДФ и фосфата и остаётся связанной с ними. (Обратимый процесс — энергия, выделившаяся в результате гидролиза, запасается в изменённой конформации миозина).Головки миозина слабо связывается со следующей субъединицей актина, фосфат отделяется, и это приводит к прочному связыванию головки миозина с актиновым филаментом. Эта реакция уже необратима.Головка претерпевает конформационное изменение, производящее подтягивание толстого филамента к Z-диску (или, что эквивалентно, свободных концов тонких филаментов друг к другу).Отделяется АДФ, за счёт этого головка отделяется от актинового филамента. Присоединяется новая молекула АТФ.

Далее цикл повторяется до уменьшения концентрации ионов Ca2+ или исчерпании запаса АТФ (в результате смерти клетки). Скорость скольжения миозина по актину ≈15 мкм/сек. В миозиновом филаменте много (около 500) молекул миозина и, следовательно, при сокращении цикл повторяется сотнями головок сразу, что и приводит к быстрому и сильному сокращению. Следует заметить, что миозиин ведёт себя как фермент — актин-зависимая АТФаза. Так как каждое повторение цикла связано с гидролизом АТФ, а следовательно, с положительным изменением свободной энергии, то процесс однонаправленный. Миозин движется по актину только в сторону плюс-конца

Источник энергии для сокращенияДля сокращения мышцы используется энергия гидролиза АТФ, но мышечная клетка имеет крайне эффективную систему регенерации запаса АТФ, так что в расслабленной и работающей мышце содержание АТФ примерно равно. Фермент фосфокреатинкиназа катализирует реакцию между АДФ и креатинфосфатом, продукты которой — АТФ и креатин. Креатинфосфат содержит больше запасённой энергии, чем АТФ. Благодаря этому механизму при вспышке активности в мышечной клетке падает содержание именно креатинфосфата, а количество универсального источника энергии — АТФ — не изменяется.

Механизм регуляцииВ основном в регуляции мышечной активности участвуют нейроны, но есть случаи, когда сокращением гладкой мускулатуры управляют и гормоны (например, адреналин и окситоцин).

Основные белки миофибрилл

ЛитератураБ. Альбертс, Д. Брей, Дж. Льюис, М. Рефф, К. Робертс, Дж. Уотсон, Молекулярная биология клетки — В 3-х т. — Пер. с англ. — Т.2. — М.: Мир, 1994. — 540 с.М. Б. Беркинблит, С. М. Глаголев, В. А. Фуралев, Общая биология — В 2-х ч. — Ч.1. — М.:МИРОС, 1999. — 224 с.: ил.











